

Растения одного вида, а иногда и таксонов более высоких рангов, обычно имеют стабильную структуру цветков (Ronse De Craene et al., 2003, 2014). Однако, у Astilbe chinensis (Maximowicz) Franche Savatier структура цветков сильно варьирует (Pan, 1995; Olson, 2000, 2001). Структура цветков может быть обусловлена предпочтениями их опылителей, так как в ходе эволюции у растений и их опылителей формируются взаимные адаптации (Timofeeff-Ressovsky, 1939; Pijl, I960; Тахтаджян, 1966; Stebbins, 1970; Fægri, 1979; Harder et al., 1996; Pellmyr et al., 1994; Pigliucci, 2003; Fenster et al., 2004; Barrett, 2014). В частности, Stebbins (2004) полагал, что в ходе эволюции наиболее частые и эффективные опылители могут обуславливать формирование специфической структуры цветков. Изменения климата могут вызывать параллельные изменения структуры цветков и их опылителей, хотя значительные расхождения также могут быть вполне ожидаемыми (Hegland et al., 2009).
Род Astilbe Buch. Ham. ex D. Don (Saxifragaceae) включает 18 видов (Pan et al., 2001) и происходит из Восточной Азии. Astilbe распространена в Восточной Азии и восточной части Северной Америки (Pan et al., 2001; Zhu et al., 2012). Согласно результатам молекулярно-филогенетического анализа, базальным таксоном в пределах рода Astilbe был японский вид A. platyphylla H. Boissieu (Kim, 1989). Дизъюнкция ареала Astilbe могла быть результатом Беренгийской миграции из Восточной Азии в Северную Америку в конце третичного периода 3,54 млн. лет (1,29-6,18 млн. лет) (Zhu et al., 2012). Среди видов Astilbe имеются виды, в популяциях которых наряду с двудомными растениями встречаются растения с двуполыми цветками (Pan et al., 2001; Olson, 2000, 2001). Astilbe chinensis, распространённая на северо-востоке Азии (Баркалов, 1987), имеет двуполые цветки, в отличие от других видов Astilbe, в популяциях которых наряду с растениями, имеющими двуполые цветки встречаются двудомные растения. В частности, к таким видам принадлежат: A. platyphylla (распространённая в Японии), A. rivularis Buchanan-Hamilton ex D. Don (распространённая в Юго-восточной Азии) и A. biternata (Ventenat) Britton ex Kearney Bull (распространённая в Северной Америке) (Kim, 1989; Pan et al., 2001; Olson, 2000, 2001). Мужские цветки у этих растений имеют двойной околоцветник (Ca5Co5A10), а женские - только чашечку (Ca5G(2)). Это сильно контрастирует с двуполыми цветками A. chinensis, которые имеют двойной околоцветник (Pan et al., 2001).
A. biternata опыляются насекомыми и ветром и образуют огромное количество нектара и пыльцы (Olson, 2000). Во время цветения А. biternata Olson (2001) выявил 30 различных видов насекомых, среди которых наиболее эффективными опылителями были Hymenoptera. Известно, что первичные опылители, которые изначально способствовали формированию адаптаций, со временем могут начать играть только второстепенные роли в репродуктивном успехе растений (Wilson et al., 1991; Waser et al., 2006). При этом насекомые, имеющие более универсальные предпочтения, могут замещать исходных опылителей, в том числе и жуков (Li et al., 2009). Исходные формы таких отношений могут быть обнаружены в рефугиумных центрах, которых не коснулись оледенения. Одним из таких центров является Приморский край России (Крестов и др., 2009), на территории которого распространена A. chinensis. Поэтому особенности опыления A. chinensis могут представлять интерес для выявления эволюционных тенденций, свойственных Astilbe.
Наши исследования были сфокусированы на анализе опылителей A. chinensis, так как прежде они не были изучены. Полиморфизм структуры цветков A. chinensis может быть обусловлен предпочтениями её опылителей, хотя в ходе эволюции их состав мог изменяться.
Исследование проводилось в период цветения на протяжении 2012-2015 гг., на территории Приморского края в естественных популяциях A. chinensis (о. Русский, окрестности Владивостока, с. Многоудобное) и коллекции Ботанического сада-института Дальневосточного отделения РАН (БСИ).
В ходе исследований была проанализирована динамика цветения, структуры побегов, соцветий и цветков у A. chinensis.
Состав визитёров A. chinensis анализировали на основании отбора проб и в ходе визуальных наблюдений. Пробы визитёров собирали в течение периода цветения A. chinensis в утренние часы (8.00-11.00), полуденные часы (12.00-15.00) и вечерние часы (16.00-19.00). Видовую принадлежность визитёров устанавливали с помощью "Определителя насекомых Дальнего Востока России" (Определитель…, 1986-1992). Параллельно проводили учёт температуры и влажности воздуха. Среди визитёров A. chinensis выявляли опылителей на основании визуальных наблюдений за поведением визитёров и по наличию пыльцы на их теле.
Для анализа полученных результатов использовали стандартные методы статистического анализа ANOVA. Для анализа зависимости визитов основных опылителей от температуры и влажности был использован метод наименьших квадратов из пакета Statistica 7.0.
Вегетационный период A. chinensis начинается в апреле, бутонизация - в июне, цветение - в июле-августе, а созревание семян - в августе-сентябре. Обычно цветение начинается в первой декаде июля. Максимальное число раскрытых цветков A. chinensis приходится на третью декаду июля и заканчивается ко второй декаде августа.
Один цветок A. chinensis цветёт в течение 3-5 суток. У A. chinensis на верхушке побегов формируется облиственный тирс (Рис. 1), образованный множеством (918,2±204,1) мелких цветков (размер: 5±1мм) (Табл. 1).

Fig. 1. The reproductive shoots of A. chinensis. A - general view; B - scheme of shoot; C - bud of the inflorescence; D - scheme of inflorescence.
Таблица 1. Структура побегов A. chinensis
Table 1. The structure of shoots in A. chinensis
| Критерии | Средняя и стандартное отклонение | Min | Max | Коэффициент вариации |
| Высота побега | 113,9±19,6 | 78,5 | 156,0 | 17 |
| Длина листьев на репродуктивном побеге | 27,1±22,4 | 58,0 | 0,5 | 78 |
| Длина соцветий | 28,9±8,4 | 15,0 | 55,0 | 29 |
| Число боковых осей на главной оси соцветия | 54,8±12,6 | 28,0 | 88,0 | 23 |
| Число цветков | 918,2±204,1 | 453,6 | 1425,6 | 22 |
| Длина цветков | 5,0±1,0 | 3,4 | 5,6 | 20 |

Fig. 2. Flowers of A. chinensis. A-C - variation of flowers structure; D - beginning of flowering.
Структура цветков A. chinensis варьирует (Табл. 1-2; Рис. 2-3). Количество пестиков имеет наиболее высокий коэффициент вариации, обычно их формируется два, но может быть от одного до трёх (Табл. 2; Рис. 2-3). Тычинки расположены в два круга. Число тычинок варьирует, во внутреннем круге сильнее, чем во внешнем круге (Табл. 2; Рис. 2-3). Тычинки имеют разную длину тычиночных нитей в пределах одного цветка и находятся на разных стадиях формирования. Длина тычиночных нитей увеличивается по мере созревания пыльников. Пестики готовы к опылению раньше, чем начинается процесс вскрывания пыльников (Рис. 2), это предотвращает самоопыление цветков.
Таблица 2. Структура цветков A. chinensis
Table 2 . The structure of flowers in A. chinensis
| Критерии | Средняя и стандартное отклонение | Min | Max | Коэффициент вариации |
| Число чашелистиков | 4,9±0,2 | 4 | 5 | 4 |
| Число лепестков | 4,9±0,2 | 4 | 6 | 4 |
| Число тычинок, | 8,4±1,8 | 4 | 14 | 21 |
| включая: | ||||
| число тычинок во внешнем круге | 5,0±0,7 | 4 | 9 | 14 |
| число тычинок во внутреннем круге | 3,4±1,5 | 0 | 5 | 44 |
| Число пестиков | 2±1,6 | 1 | 3 | 80 |
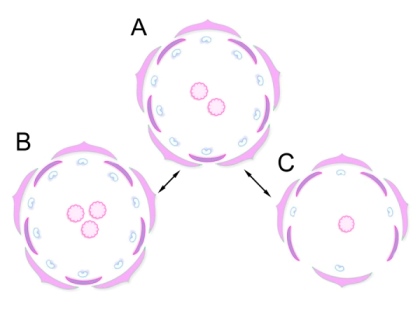
Fig. 3. Diagrams of flowers in A. chinensis.
Наши исследования показали, что визитёры A. chinensis имели разную частоту встречаемости и динамику посещений (Табл. 3-4, Рис. 4-6). Insecta и Araneae были основными визитерами A. chinensis (Рис. 4-5). В числе Insecta обычно встречались Coleoptera, Hymenoptera, Heteroptera и Diptera (Рис. 5-6). Coleoptera преобладали в начале и середине цветения, а Hymenoptera в середине и в конце цветения (Рис. 6 A). Coleoptera чаще всего посещали цветки в утренние и вечерние часы, а Hymenoptera в полуденные (Рис. 6 B). Heteroptera и Coleoptera имели сходную динамику посещения цветков A. chinensis, но Heteroptera являются более редким визитёрами (Рис. 6 A). Таким образом, доминирующие визитёры A. chinensis имели разные предпочтения для посещений (Рис. 4). Исследования показали, что температура и влажность воздуха имели достоверное влияние на посещения Coleoptera (коэффициенты корреляции rt=-0,88, rh=0,63) и Hymenoptera (rt=-0,13, rh=-0,83) (Рис. 7). Coleoptera реже всего появляются при низкой температуре и низкой влажности в рамках измеренных значений, зато, когда температура соответствуют 20° С, а влажность - 80 мм, присутствие Coleoptera становится постоянным (Рис. 7 A). Визиты Hymenoptera в значительной степени зависели от сочетания температуры и влажности. Обилие Hymenoptera увеличивается по мере возрастания температуры, причем сочетание температуры и влажности так же имеет значение (Рис. 7 В). Таким образом, температура и влажность являются факторами, лимитирующими посещения Coleoptera и Hymenoptera цветков A. chinensis. При этом Coleoptera имеют более жесткое ограничение чем Hymenoptera. В целом, Hymenoptera предпочитают более аридные условия для посещения цветков A. chinensis чем Coleoptera.

Fig. 4. Visitors of flowers in A. chinensis. A-B - Anoncodes W. Schmidt (Oedemeridae); C - Sipalus L. (Curculionidae); D - Apis L. (Apidae).
Таблица 3. Посетители цветков A. chinensis
Table 3. Flowers visitors of A. chinensis
| Класс | Отряд | Семейство | 1 | 2 | 3 | 4 |
| Insecta L.
|
Coleoptera L. | Bostrichidae Latrelle | + | - | - | + |
| Cerambycidae Latrelle | + | - | - | - | ||
| Chrysomelidae Latrelle | + | + | + | + | ||
| Coccinellidae Latrelle | + | + | + | + | ||
| Curculionidae Latrelle | + | + | + | + | ||
| Mordellidae Latrelle | + | + | + | + | ||
| Oedemeridae Latrelle | + | + | - | - | ||
| Scarabaeidae Latrelle | + | + | + | + | ||
| Diptera L. | Culicidae | + | - | - | - | |
| Drosophilidae Rododani | + | - | - | - | ||
| Entomophthora muscae (Cohn) Fresenius | + | + | + | - | ||
| Scathophagidae | + | + | + | + | ||
| Simuliidae | + | + | + | + | ||
| Syrphydae Rondani | + | + | + | + | ||
| Hemiptera L. | Aphidoidea Latrelle | + | + | + | + | |
| Cicadellidae Latrelle | + | - | - | + | ||
| Coreidae Amyot and Serville | + | + | + | + | ||
| Miridae Hahn. | + | + | + | + | ||
| Pentatomidae Leach | + | + | + | - | ||
| Rhopalidae Amyot and Serville | + | - | - | - | ||
| Hymenoptera L. | Apidae | + | + | + | + | |
| Formicidae Latrelle | + | + | + | + | ||
| Ichneumonidae Latrelle | + | + | + | + | ||
| Lepidoptera L. | Papilionoidea Latrelle | + | + | + | + | |
| Macroheterocera | + | + | + | + | ||
| Orthoptera Latrelle | Acrididae MacLeay | + | + | - | - | |
| Thysanoptera Halday | Melanthripidae Bagnall | - | + | + | + | |
| Arachnida Cuvier. | Arneae Clerck | Thomisidae Sundevall | + | + | + | + |
Примечание: 1. БСИ; 2. о. Русский; 3. окрестности г. Владивосток (пригород); 4. окрестности с. Многоудобное Приморского края.
Notates: 1. Botanical Garden-Institute; 2. is. Russkiy; 3. The neighborhood of Vladivostok (Prygorod); 4. Mnogoudobnoe, from Primorsky kray.
Состав визитёров A. chinensis в природных популяциях и в коллекции отличался (Табл. 3). Возможно, что это связано с разницей фонового окружения. Из представителей Coleoptera на цветках A. chinensis чаще всего встречались Oedemeridae, Mordellidae, Scarabaeidae и Curculionidae (Табл. 3). Они имеют крошечные размеры (3±15 мм, 0,03±0,07 г) и могут длительное время остаться на цветах. Пыльца остаётся между волосками на их теле, и они переносят её на другие цветки. Среди Hymenoptera чаще всего встречались Formicidae и Apidae (Рис. 6), но они имели разную динамику посещений. Apidae чаще встречались в полдень, а Formicidae утром и вечером (Рис. 6). Apidae были основными опылителями среди Hymenoptera, их размеры (16±9 мм, 0,44±0.3 г) были крупнее чем у Coleoptera, посещавших цветки A. chinensis. Кроме того, Apidae влетали в цветок на большой скорости, в результате чего побег начинал раскачиваться как маятник. Таким образом, при опылении цветков A. chinensis, Apidae создавали явно большую механическую нагрузку, чем Coleoptera.
Посетители A. chinensis (Coleoptera) откладывают личинки в завязи цветка, то есть являются паразитами этого растения.
Formicidae (Hymenoptera) посещали A. chinensis после начала массового цветения, при этом их визиты приходились в основном на утренние или вечерние часы. В этот период на A. chinensis также появилась Aphidoidea. Возможно, что с этим фактом и были связаны визиты Formicidae на цветки A. chinensis.
Во второй половине периода цветения A. chinensis на растениях появлялись Arachnida. Они предпочитали утренние и дневные часы для своих визитов. Выбор времени для визитов Arachnida был связан с оптимальным периодом для их охоты. В частности, к моменту их появления в цветках A. chinensis появлялись Thysanoptera, на которых охотились Arachnida. Учитывая, что в это время начинали развиваться семена, то Thysanoptera представляют угрозу для их развития. Поэтому визиты Arachnida могли способствовать очистке цветков от вредных насекомых, то есть они выполняют роль «санитаров».
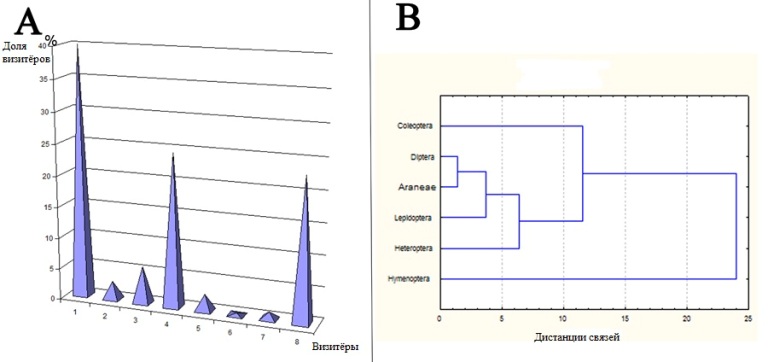
Рис. 5. Распределение посетителей цветков A. chinensis. A - долевое отношение: 1 - Coleoptera, 2 - Diptera, 3 - Heteroptera, 4 - Hymenoptera, 5 - Lepidoptera, 6 - Orthoptera, 7 - Thysanoptera, 8 - Araneae; B - распределение насекомых по частоте посещений.
Fig. 5. The distribution of flowers visitors in A. chinensis. A - ratio of shares: 1 - Coleoptera, 2 - Diptera, 3 - Heteroptera, 4 - Hymenoptera, 5 - Lepidoptera, 6 - Orthoptera, 7 - Thysanoptera, 8 - Araneae. B - distribution in reation to visits frequency of insect groups.
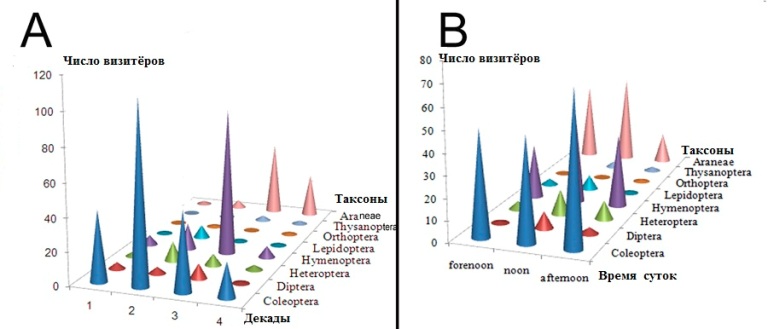
Рис. 6. Динамика посещения цветков A. chinensis. A - Динамика посещения цветков на протяжении периода цветения; B - Динамика посещения цветков в течение дня.
Fig. 6. Dynamics of visits flowers in A. chinensis. A - Dynamics of visits during flowering time. B - Dynamics of visits during day.
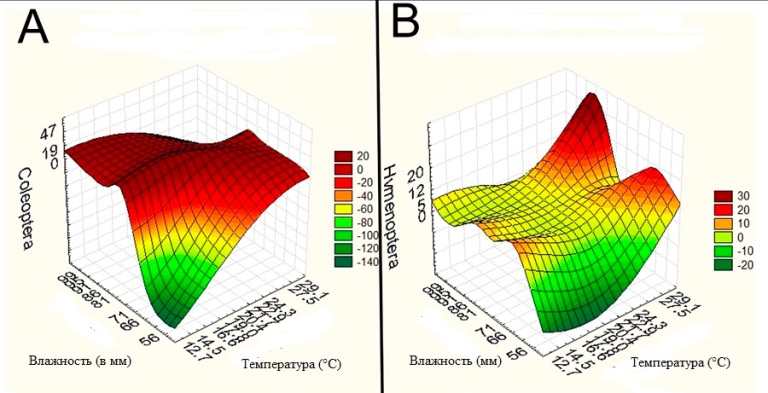
Рис. 7. Влияние температуры и влажности на визиты основных опылителей A. chinensis. A - Влияние Effects of temperature and humidity on visits of Coleoptera; B - Effects of temperature and humidity on visits of Hymenoptera.
Fig. 7. Effects of temperature and humidity on visits of main pollinators of A. chinensis. A - Effects of temperature and humidity on visits of Coleoptera; B - Effects of temperature and humidity on visits of Hymenoptera.
Исследования показали, что A. chinensis формирует соцветия из множества мелких цветков с варьирующей структурой, формирующих большое количество пыльцы в течение относительно продолжительного периода, это способствует привлечению широкого спектра визитёров, имеющих разные предпочтения для своих визитов на растения. Чаще всего на цветках A. chinensis встречались Coleoptera, Hymenoptera, Heteroptera и Arachnida. Основные опылители принадлежали к Coleoptera и Hymenoptera.
Температура и влажность воздуха влияют на состав опылителей в период цветения. Hymenoptera предпочитали посещать цветки при более высоких температурах и более низкой влажности чем Coleoptera.
В числе опылителей A. chinensis были обнаружены Coleoptera, которые не были отмечены в числе опылителей Astilbe прежде. Это является поводом для пересмотра характеристик растений опыляемых жуками. Большое количество пыльцы, по-видимому, является решающим фактором при опылении жуками, а размеры околоцветника значения не имеют.
Место жуков в эволюции опыления Astilbe представляет значительный интерес, так как жуки были в числе первых опылителей базальных Angiospermae (Diels, 1916; Гринфельд и др., 1958; Тахтаджян, 1966; Pijl, I960; Baker, 1955 Красилов и др., 1989), хотя и не единственными (Малышев, 1966; Krassilov et al., 2008). Опыление жуками связывают с рядом структурно-функциональных адаптаций. В частности, если растения опыляются жуками, то они нуждаются в крупном цветке и кортикальной проводящей системе (обеспечивающей устойчивость «посадочной площадки» при посадке относительно массивных насекомых) (Тахтаджян, 1966; Krassilov et al., 2014). Эти характеристики соответствуют морфологии цветков Magnolia и Nelumbo, но они совсем не подходят для Astilbe. Это является поводом для пересмотра признаков, характерных для карантофильных растений. Результаты исследования показали, что Coleoptera могут создавать гораздо меньшую механическую нагрузку чем Hymenoptera, поэтому развитие кортикальной проводящей системы для кантарафильных растений вряд ли необходимо.
Наличие крупного околоцветника принадлежит к числу признаков, характерных для кантарофильных растений. Однако Coleoptera опыляют как растения с крупными цветками (Magnolia, Nelumbo) (Borsch et al., 1994; Mathews et al., 2000), так и Cycadales Pers. ex Bercht. & J. Presl (Hall et al., 2011; Yu, 2015), у которых отсутствует не только околоцветник, но и сами цветки. Quasada et al. (2006) обратили внимание на большое количество пыльцы, свойственное растениям, которые опыляются жуками. В частности, в качестве примера он приводит представителей Araceae, которые имеют голые цветки. Поэтому тот факт, что соцветия A. chinensis формируют много пыльцы, мог иметь решающее значение для её опылителей. Возможно, что размер околоцветника не имеет значения для опыления жуками, а решающее значение имеет значительный объём пыльцы.
Мы выражаем искреннюю признательность профессору А. С. Лелей и старшему научному сотруднику С. А. Шабалину из Биолого-почвенного института ДВО РАН, а также старшему научному сотруднику Е. В. Конюховой из Зоологического музея Дальневосточного федерального университета за помощь при идентификации таксонов насекомых, которые были обнаружены в процессе исследований.
References
Barkalov V. Yu. Saxifragaceae. Sosudistye rasteniya Sovetskogo Dalnego Vostoka Vascular plants of the Soviet Far East / Otv. red. S. S. Kharkevitch. L.: Nauka, 1987. T. 2. S. 393—397.
Grinfeld E. K., Issi I. V. Rol zhukov v opylenii rastenij The role of adults (beetles) in the pollination of plants // Utchenye zapiski LGU. 1958. № 240. Seriya biologitcheskikh nauk. № 46. S. 148—159.
Krasilov V. A. Proiskhozhdenie i evolyutsiya tsvetkovykh rastenij. The origin and early evolution of flowering plants M.: Nauka, 1989. 264 s.
Krestov P. V., Barkalov V. Yu., Omelko A. M., Yakubov V. V., Nakamura Yu., Sato K. Reliktovye kompleksy rastitelnosti sovremennykh refugiumov severo-vostotchnoj Azii Relic vegetation complexes in the modern refugia of Northeast Asia // Komarovskie tchteniya. Vladivostok, 2009. № 56. S. 5—63.
Malyshev S. I. Stanovlenie perepontchatokrylykh i fazy ikh evolyutsii. Formation of Hymenoptera and phase of their evolution M.–L.: Nauka, 1966. 326 s.
Opredelitel nasekomykh Dalnego Vostoka Rossii Key to the insects of Soviet Far East of Russia / Pod red. A. S. Lelej. L.: Nauka, 1986—1992.
Takhtadzhyan A. L. Sistema i filogeniya tsvetkovykh rastenij. System and Phylogeny of Flowering Plants M.-L.: Nauka, 1966. 611 s.
Baker H. G. Self-compatibility and establishment after “long distance” dispersal // Evolution. 1955. Vol. 9. P. 347—348.
Barrett S. C. H. Evolution of mating systems: outcrossing versus selfing. In The Princeton Guide to Evolution Edited by Losos J. New Jersey. Princeton University Press. Princeton: 2014. P. 356—362.
Borsch T., Barthlott W. Classification and distribution of the genus Nelumbo Adans. (Nelumbonaceae) // Beiträge zur Biologie der Pflanzen. 1994. Vol. 68. P. 421—450.
Diels L. Käferblumen bei Ranales und ihre Bedeutung für die Phylogenie der Angiospermen // Ber. Deutsch. Bot. Ges. 1916. Vol. 34. P. 758—774.
Fægri K., van der Pijl L. The principles of pollination ecology. 3rd edn. Oxford: Pergamon Press, 1979. 244 p.
Fenster C. B., Armbruster W. S., Wilson P., Dudash M. R., Thomson J. D. Pollination syndromes and floral specialization. Annual Review of Ecology // Evolution and Systematics. 2004. Vol. 35. P. 375—403.
Hall J. A., Walter G. H. Does pollen aerodynamics correlate with pollination vector? Pollen settling velocity as a test for wind versus insect pollination among Cycads (Gymnospermae: Cycadaceae: Zamiaceae) // Biological journal of the Linnean society. 2011. Vol. 104. № 1. P. 75—92.
Harder L. D., Barrett S. C. H. Pollen dispersal and mating patterns in animal-pollinated plants / In: Floral biology: studies on floral evolution in animal-pollinated plants / Edited by Lloyd D. G., Barrett S. C. H. New York: Chapman and Hall, 1996. P. 140–190.
Hegland S. J., Nielsen A., Lázaro A., Bjerknes A. L., Totland O. How does climate warming affect plant-pollinator interactions? // Ecology Letters. 2009. Vol. 12. P. 184—195.
Krassilov V. A., Barinova S. S. “Flower” of Magnolia grandiflora is not flower and what about “basal angiosperms” // Journal of Plant Sciences. 2014. Vol. 2. № 6. P. 282—293.
Krassilov V. A., Volynets E. B. Weedy Albian angiosperms // Acta Palaeobotanica. 2008. Vol. 48. № 2. P. 151–169.
Kim Y. D. Molecular phylogeny of Astilbe: Implications for phylogeography and morphological evolution // Korean journal of plant taxonomy. 2009. Vol. 39. № 1. P. 35—41.
Li J-K., Huang S. Q. Effective pollinators of Asian sacred lotus (Nelumbo nucifera): Contemporary pollinators may not reflect the historical pollination syndrome // Annals of botany. 2009. Vol. 104. № 5. P. 845—851.
Mathews S., Donoghue M. J. Basal angiosperm phylogeny inferred from duplicate phytochromes A and C Int // J. Plant Sci. 2000. Vol. 161. № 6. P. 41—55.
Olson M. S., Antonovics J. Correlation between male and female reproduction in the subdioecious herb Astilbe biternata (Saxifragaceae) // American Journal of Botany. 2000. Vol. 87. № 6. P. 837—844.
Olson M. S. Patterns of fruit production in the subdioecious plant Astilbe biternata (Saxifragaceae) // Journal of Ecology. 2001. Vol. 89. № 4. R. 600—607.
Pan J. T. A study of the tribe Astilbeae Miq. (Saxifragaceae) // Acta Phytotaxonomica Sinica. 1995. Vol. 33. P. 390—402.
Pan J. T., Ohba H. Astilbe / In Flora of China. St. Louis: Sci. Press. Beijing. Miss. Bot. Gard. Press. 2001. Vol. 8. P. 274—276.
Pellmyr O., Huth C. J. Evolutionary stability of mutualism between yuccas and yucca moths // Nature. 1994. Vol. 372. P. 257—260.
Pigliucci M. Phenotypic integration: studying the ecology and evolution of complex phenotypes // Ecology Letters. 2003. Vol. 6. P. 265—272.
Pijl L. Ecological aspects of flower evolution // Evolution. I960. Vol. 14. № 15. P. 403—416.
Quesada M., Rosas F., Herrerias-Diego Y., Aguliar R., Lobo J. A., Sanchez-Montoya G. Evolutionary Ecology of Pollination and Reproduction of Tropical Plants // Tropical biology and conservation management. 2006. P. 5.
Ronse De Craene L. P., Iwamoto A., Bull-Hereñu K., Dos Santos P., Luna J. A. Understanding the structure of flowers. The wonderful tool of floral formulae: A response to Prenner & al. // Taxon. 2014. Vol. 63. № 5. P. 1103—1111.
Ronse De Craene L. P., Soltis P. S., Soltis D. E. Evolution of floral structures in basal angiosperms // International Journal of Plant Science. 2003. Vol. 164. P. 329—363.
Stebbins G. L. Adaptive radiation of reproductive characteristics in angiosperms. I: Pollination mechanisms // Annual Review of Ecology and Systematics. 1970. Vol. 1. P. 307—326.
Timofeeff-Ressovsky N. W. Genetik und Evolution (Bericht einer Zoologen) / Z. Indukt. Abstammungs-Vererbungsl. 1939. Vol. 76. P. 158—218.
Waser N. M., Ollerton J. Plant-pollinator interactions // From specialization to generalization. Chicago: University of Chicago Press. 2006.
Wilson P., Thomson J. D. Heterogeneity among floral visitors leads to discordance between removal and deposition of pollen // Ecology. 1991. Vol. 72. P. 1503—1507.
Yu W-B. Floral and reproductive ecology of Cycas panzhihuaensis // ATBC Asia Pacific chapter meeting, at Phnom Penh conference paper. March, 2015.
Zhu W-D., Nie Z-L., Jun W., Sun H. Molecular phylogeny and biogeography of Astilbe (Saxifragaceae) in Asia and eastern North America // Botanical Journal of the Linnean Society. 2012. Vol. 171. P. 377—394.
