

Введение
Раковинные амебы (Testacea) распространены повсеместно. Их можно обнаружить в водоемах, болотах, влажной почве, сфагновых мхах. В биоценозах они выполняют функцию минерализации органических веществ, делая последние доступными для других микроорганизмов и растений. Особую значимость деятельность тестацей приобретает в верхних слоях почвы ввиду того, что они участвуют в преобразовании органических соединений в гумус (Гельцер и др., 1980). В экосистемах раковинные амебы в основном являются сапрофитной группой, но встречаются и хищные виды. Например, представители родов Euglypha и Difflugia регулируют численность микроорганизмов в почве. В ходе исследований у них был выявлен разнообразный рацион питания, который включал коловраток, инфузорий, динофлагеллят, мелкие почвенные частицы; отмечен также каннибализм (Bo-Ping Han et al., 2011).
Видовой состав тестацей является индикатором физико-химических свойств воды и почвы, что, в свою очередь, может находить применение в биодиагностике и анализе изменяющихся условий окружающей среды (Robert, Ellison, 1995). Его используют в проверке торфяных залежей для выявления горизонтов с различной степенью обводнения (Мазей, Цыганов, 2006). Кроме того, среди методов палеоэкологических реконструкций известен ризоподный анализ, который помогает реконструировать поверхностную влажность болот на основе состава раковинных амеб в торфе (Charman et al., 2000; Chambers et al., 2010; Цыганов и др., 2020).
Раковинные амебы обладают высокой чувствительностью к изменениям внешних условий. Например, их используют в качестве индикаторов поверхностной влажности болот и состояния пресноводных экосистем (Цыганов и др., 2020). Почвенные виды тестацей чувствительны и к антропогенному воздействию. Например, в почвах г. Томска в местах с высокой нагрузкой (вблизи ГРЭС, дорог или застроек) отмечено уменьшение количественного состава тестацей, в отличие от мест с минимальной нагрузкой (естественные лесные ценозы) (Кулюкина, 2015).
Почвы садов и парков также испытывают антропогенное воздействие, что негативно сказывается на состоянии обитающих здесь различных групп организмов. В Ботаническом саду ПетрГУ, где проводились исследования, воздействие на почву умеренное. Сад был создан в середине прошлого столетия с научными целями, а также как флористический питомник ― для озеленения городов и поселков Республики Карелия. Территория сада обеспечивает проведение научно-исследовательской, просветительской и учебных практик.
Исследования, посвященные составу сообществ раковинных амеб в биоценозах Карелии, малочисленны. Имеются лишь сведения по видовому составу раковинных амеб в Лоухском районе (выявлено 24 вида). Авторами установлено и соответствие видового состава разным уровням увлажнения и кислотности в осоково-сфагновом лесу (Мазей, Кабанов, 2008). По северу известны исследования тестацей олиготрофных болот Онежского района (Архангельская обл.) (Кац, 1971).
Целью настоящей работы являлось изучение сообщества раковинных амеб в типичных для Республики Карелия биоценозах. Были рассмотрены почвенные виды тестацей в сосновых биоценозах и на лугу, расположенных на территории Ботанического сада ПетрГУ. Данная территория характеризуется минимальным антропогенным воздействием, так как находится в охраняемой зоне, что, в свою очередь, способствовало получению наиболее точных результатов.
Объекты и методы исследований
Исследования проводили в Ботаническом саду ПетрГУ в двух типах биотопов: злаково-разнотравный луг и сосняк кисличный. Период исследований ― с 2019 по 2020 г. Почвенные пробы собирали в теплый период (май ― октябрь) на четырех площадках (рис. 1).
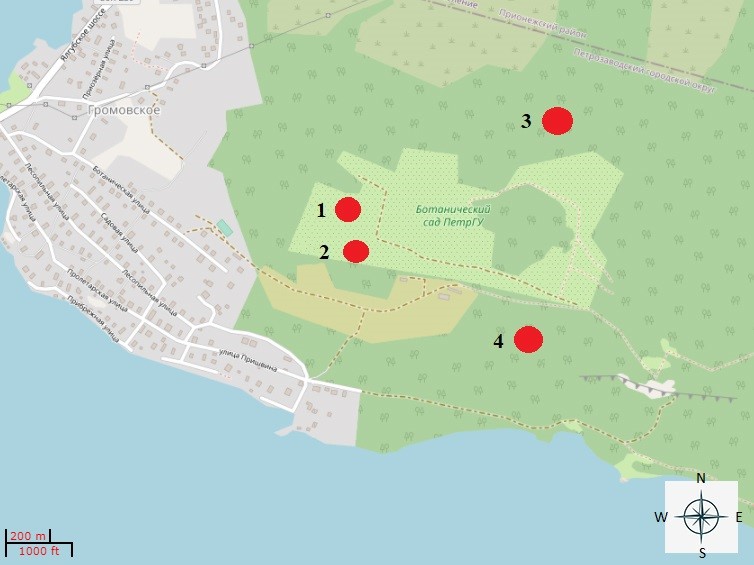
Рис. 1. Места отбора проб почвенных образцов на территории Ботанического сада ПетрГУ(цифрами отмечены исследуемые площадки): 1, 2 ― луг злаково-разнотравный; 3, 4 ― сосняк кисличный (подсекция сосняки мелкотравно-зеленомошные).
Для почв сосняка кисличного характерны грубогумусовые супесчаные бурозёмы. Отличительной чертой данной территории является наличие гумус-продуктивного горизонта, где происходит гумификация органических веществ. Результаты химических анализов показали, что почвы имеют характерную для таежной зоны сильнокислую реакцию среды и высокую гидролитическую кислотность, особенно в подстилке (Красильников, Платонова, 2001).
Почва злаково-разнотравного луга представлена торфяно-глеезёмом на флювиогляциальных песках, для которых характерна слабокислая и близкая к нейтральной реакция среды, а также высокое содержание магния и кальция (Красильников, Платонова, 2001).
При изучении видового состава раковинных амеб пробы отбирали с четырех исследуемых площадок: по две пробы из каждого изучаемого биотопа (рис. 1). Все образцы почв изымали из гумусового горизонта, что позволило исследовать сообщества раковинных амеб, обитающих на глубине до 10–20 см. Для выявления сезонной динамики образцы почвенных проб с одной площадки собирали ежемесячно с мая по октябрь в течение двухлетнего периода.
Из почвенной пробы раковинных амеб выделяли по методике А. А. Рахлеевой, Г. А. Коргановой (2005). В работе использовались равные по весу (5 г) образцы почв с частицами детрита, которые помещали в колбу, заливали произвольным количеством воды (150―200 мл) и оставляли на несколько часов для размокания почвенных частиц. Далее взвесь взбалтывали в течение 10 мин и фильтровали через сито с ячеями 0,8 мм, после чего отстаивали в течение 2 ч. Образующуюся надосадочную жидкость сливали, а оставшееся количество фильтрата переносили в градуированную емкость и снова давали отстояться. Суспензию окрашивали кармином и фиксировали формалином в течение суток. Живые клетки простейших и цисты приобретали розовый цвет, что позволяло регистрировать их как активные организмы. С каждой пробы просматривалось не менее десяти препаратов.
Подсчет раковинных амеб проводили в водных суспензиях при объеме ~45 мкл. Каплю подготовленного раствора помещали на предметное стекло с помощью пипетки Пастера. При определении численности учитывали как живых особей, так и пустые раковинки, которые обычно составляют значительную часть сообществ тестацей. Микроскопирование препаратов проводилось при увеличении х100 и х65 с использованием микроскопов Axio Scope A1 и Motic 1500. Всего было изучено более 1400 препаратов. Проверка правильности определения видов выполнена кандидатом биологических наук Курьиной И. В. (Институт мониторинга климатических и экологических систем СО РАН, г. Томск).
Анализ данных
Для оценки достоверности различий в видовом богатстве и приуроченности вида к биотопу использовали критерий Пирсона (χ2). Значимыми принимали различия при условии р < 0.05. При изучении разнообразия сообщества раковинных амеб применяли индекс доминирования (С), позволяющий оценить степень равномерности распределения признаков объектов выборки, а высокие значения выявляли преобладание численности определенных видов в изучаемой местности (Коросов, 2007).
Сходство между почвенными пробами устанавливали с помощью коэффициента Чекановского ― Сёренсена. Коэффициент показывает близость или отдаленность биотопов по количественному составу выявленных видов. При вычисленном значении, равном 100 %, объясняется полное совпадение видов на двух участках, а при нуле ― абсолютной несхожести и отсутствии хотя бы одного общего вида (Песенко, 1982).
Результаты и обсуждение
В исследованных биотопах проведен учет семи таксонов раковинных амеб (см. табл.). Наибольшее количество раковинок отмечено для видов Cyclopyxis eurystoma и Trinema lineare.
Вид C. eurystoma имеет раковину средних размеров, в профиль полусферической формы, диаметром 30―80 мкм и высотой 30―50 мкм (рис. 2а). Устье C. eurystoma округлое, диаметром 22―34 мкм. Покрывающие раковинки минеральные частицы крупные и хорошо заметны.
Вид T. lineare характеризуется мелкой удлиненно-яйцевидной раковинкой. Он хорошо различим и по округлому устью диаметром до 10 мкм (рис. 2б). Его раковинки небольших размеров, длиной 16―35 мкм, шириной 7―17 мкм. В биоценозах T. lineare играет важную роль, расщепляя лигнин, тем самым способствуя образованию органоминеральных компонентов почвы (Корганова, 1997).
Таблица. Встречаемость почвенных раковинных амеб (Testacea) в биотопах исследования. Значения показателей критерия Пирсона (χ2) и уровня значимости (p) при числе степеней свободы df = 1
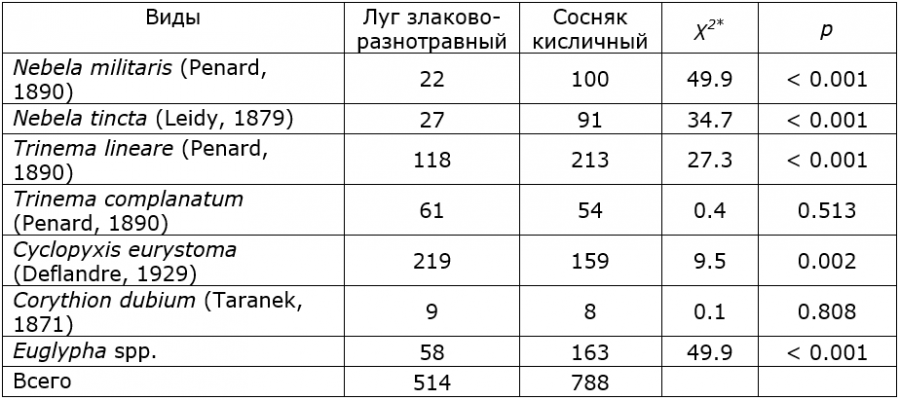

Рис. 2. Доминирующие виды раковинных амеб в почвах Ботанического сада ПетрГУ:a ― Cyclopyxis eurystoma (Deflandre, 1929); b ― Trinema lineare (Penard, 1890).
Раковинка Nebela militaris имеет узкогрушевидную форму средних размеров (длина 50―80 мкм, ширина 25―50 мкм), которая сужается равномерно к устью, а в приустьевой части слегка расширяется (рис. 3а). Раковинка покрыта круглыми широкоэллиптическими палочковидными или неправильной формы идиосомами. Устье эллиптическое, шириной 16―20 мкм, в профиль вырезанное. У Nebela tincta раковинка относительно крупная, грушевидная или яйцевидная, латерально-уплощенная (длина 75―95 мкм, ширина 55―64 мкм) (рис. 3б). Чаще всего раковина имеет светло-желтый оттенок, а мелкие идиосомы иногда покрывают только ее часть. Устье эллиптическое, шириной 20 мкм; по бокам имеются две не всегда четко выраженные дополнительные поры.
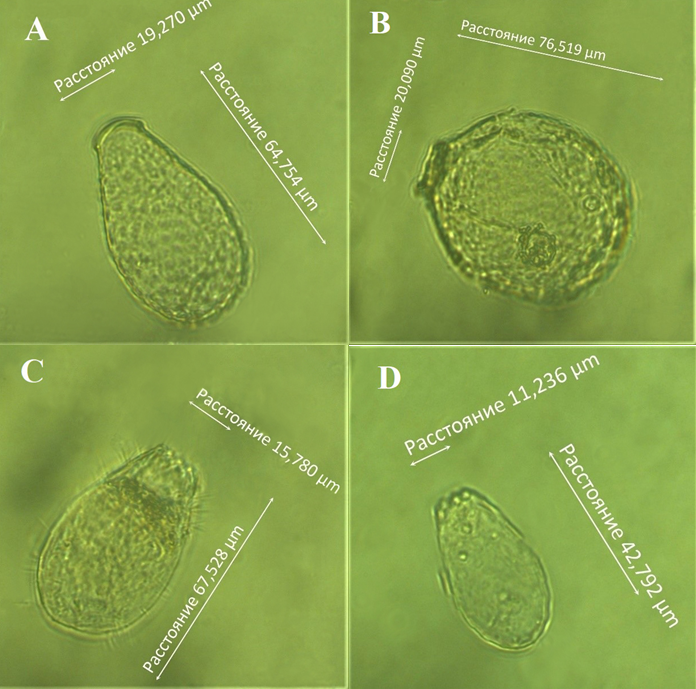
Рис. 3. Представители раковинных амеб: a ― Nebela militaris (Penard, 1890); b ― Nebela tincta (Leidy, 1879); c, d ― виды рода Euglypha.
Наиболее часто в пробах встречались представители рода Euglypha. Эти виды довольно крупных размеров (длина 40―70 мкм, ширина более 25 мкм) (см. рис. 3в, г). Раковина состоит из перекрывающихся пластинок (идиосом), обычно расположенных продольными рядами. Сами идиосомы имеют зубчатую форму и обрамляют устье раковины. Некоторые авторы отмечают, что морфология раковинок у этого рода очень изменчива и зависит от местообитания. Например, виды, которые встречаются на влажных участках, на раковинке имеют многочисленные тонкие иглы, а у видов, обитающих в сухих условиях, иглы могут отсутствовать (Bobrov et al., 2002).
Выявленные виды раковинных амеб были отмечены во всех исследуемых пробных площадках, но количество их было неодинаково. Для большинства тестацей установлено биотопическое предпочтение. В сосняках чаще встречались представители родов Nebela, Euglypha и Trinema lineare (см. табл.). Последний вид также широко распространен в сосновых лесах в Среднем Поволжье, Центральной и Восточной Сибири (Смолянинова, Гренадерова, 2018; Булатова, 2010; Мазей, Ембулаева, 2009). Выявленные нами виды рода Nebela часто встречаются и в почвах сосняков (Кулюкина и др., 2016). Кроме того, известно, что большинство видов рода Nebela обитает в болотах (Мазей и др., 2009).
Для разнотравного луга характерен вид Cyclopyxis eurystoma. Вычисленное эмпирическое распределение отличается достоверно (см. табл.). В других источниках указано, что C. eurystoma является эврибионтом (Мазей, Комаров, 2015).
Неподходящими условиями для обитания Corythion dubium оказались почвы луга и сосняка. В данных биотопах вид отмечен в незначительном количестве (см. табл.). Известно, что C. dubium часто встречается в осоково-сфагновом заболоченном лесу, оставаясь одним из доминирующих в сообществе (Мазей, Кабанов, 2008). Он также обнаруживается на эпилитных мхах (Белякова, 2010).
Индекс доминирования (С) сообщества раковинных амеб в почвах луга выше, чем в ельниках, и составляет 0.26. В открытых биоценозах среди отмеченных видов тестацей преобладает лишь C. eurystoma (см. табл.). В сосняке кисличном структура населения более выравнена, поэтому индекс доминирования имеет меньшее значение и составляет 0.19.
Значение коэффициента сходства биотопов Чекановского — Сёренсена позволило выделить два кластера сообщества раковинных амеб: луговой и сосновый, с общностью между ними 73 % (рис. 4). Сходство проб соснового биоценоза (пробы 3 и 4) составляет 91 %, а лугового (пробы 1 и 2) — 83 %.
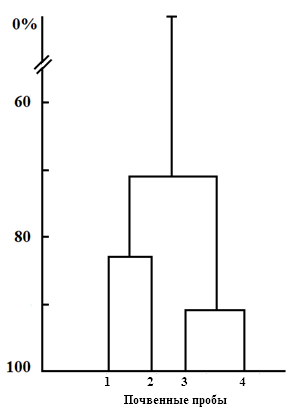
Рис. 4. Сходство обилия видов тестацей в исследуемых пробных площадках злаково-разнотравного луга (1, 2) и сосняка кисличного (3, 4).
Сезонная динамика
Для большинства выявленных видов наблюдается изменение численности во время вегетационного периода (рис. 5). С мая по август регистрируется увеличение числа раковинных амеб в пробах, а с августа по сентябрь — eго спад. В течение всего весенне-осеннего периода наибольшее количество C. eurystoma и T. lineare выявлено в июле, а N. militaris за все время наблюдений встречался равномерно. Однако не исключено, что в течение сезона может наблюдаться перемещение раковинных амеб в почвенных горизонтах. Такое изменение происходит, как правило, за счет токов воды (Рахлеева и др., 2011). Качественный состав тестацей изменяется незначительно в течение весенне-осеннего сезона (Трулова, Мазей, 2012).
По сравнению с болотными биоценозами лесные не отличаются большим видовым разнообразием раковинных амеб ввиду меньшей увлажненности. Например, в болотных экосистемах верховья р. Суры выявлено 59 видов и форм ризопод (Мазей и др., 2007). В почвах лесов общее число видов тестацей значительно меньше: на территории биоценозов южной тайги (Центрально ― Лесной государственный природный заповедник, Тверская область) зарегистрировано 24 вида (Рахлеева, Корганова, 2005); в почвенном покрове северотаежных лиственничников (Эвенкийский р-он, Красноярский край, п. Тура) был обнаружен 31 вид раковинных амеб (Безкоровайная и др., 2017).
Основным фактором, определяющим дифференциацию сообществ раковинных амеб, является влажность субстрата (Бобров и др., 2002). В болотах на территории Украины видовое разнообразие сообщества раковинных амеб обнаруживается в наиболее увлажненных условиях, и с глубиной (до 16 см) как видовое богатство (с 8 до 27 видов), так и численность (от 14 до 74 тыс. экз. в г почвы) раковинных амеб постепенно возрастает (Бубнова, Мазей, 2008).
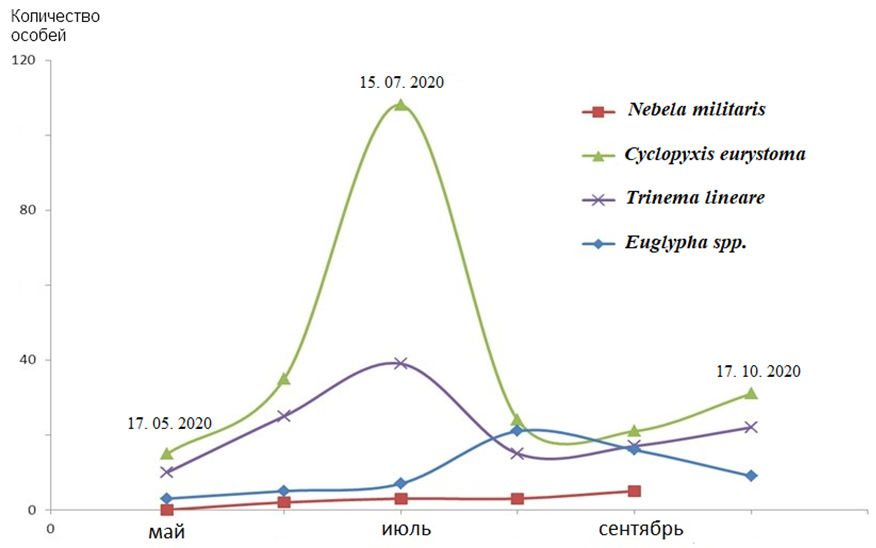
Рис. 5. Сезонная динамика численности почвенных видов тестацей злаково-разнотравного луга.
Для выявленных видов в Ботаническом саду ПетрГУ определено сходство по местообитаниям в других регионах. Виды C. eurystoma и T. lineare являются доминирующими в сообществе, их высокая численность отмечена в сосновых, смешанных лесах и в болотах биоценозов южной тайги (Булатова, 2010; Мазей, Бубнова и др., 2009; Бабешко и др., 2015; Цыганов и др., 2020). На протяжении всего вегетационного сезона данные виды остаются преобладающими и в подстилке сосняков Юксеевского лесничества (северная граница Красноярской лесостепи, Большемуртинский р-он, Красноярский край) (Смолянинова, Гренадерова, 2018). Также вид T. lineare обнаруживается более чем в 80 % всех образцов, взятых с территории болотных экосистем парка «Орловское Полесье» (северо-запад Орловской области, восточная периферия Калужско-Брянского лесного массива) (Бабешко и др., 2015).
Благодарности
Авторы выражают благодарность Анжелле Валерьевне Сониной, заведующей кафедрой ботаники и физиологии растений ПетрГУ, за возможность проведения лабораторных работ и Курьиной И. В., старшему научному сотруднику Института мониторинга климатических и экологических систем СО РАН (г. Томск), за помощь в определении материала и работе над рукописью.
Литература
Белякова О. И. Структура сообществ раковинных амеб и гетеротрофных жгутиконосцев в эпифитных и эпилитных мхах и лишайниках (автореферат) , Саратов: Саратовский государственный университет им. Н. Г. Чернышевского, 2010. 24 с.
Бобров А. А., Чармен Д., Уорнер Б. Экология раковинных амеб олиготрофных болот (особенности биологии политипических и полиморфных видов) // Известия РАН. Сер. биологическая. 2002. № 6. С. 738—751.
Бубнова О. А., Мазей Ю. А. Структура сообщества раковинных амеб (Testacealobosea; Testaceafilosea; Amphitremidae) в сфагновых биотопах Романовского болота (Киевское Полесье) // Известия ПГПУ им. В. Г. Белинского. 2008. № 10 (14). С. 88—93.
Булатова У. А. Фауна и экология раковинных амёб (Rhizopoda, Testacea) сосновых лесов Томской и Кемеровской областей // Вестник Томского государственного университета. Биология. 2010. № 2 (10). С. 58—67.
Безкоровайная И. Н., Климченко А. В., Гренадерова А. В. Влияние лесных пожаров на биоту криогенных почв // Теоретические и прикладные аспекты лесного почвоведения: Сборник материалов VII Всероссийской научной конференции по лесному почвоведению с международным участием . Петрозаводск: Карельский научный центр РАН, 2017. C. 182—185.
Гельцер Ю. Г., Корганова Г. А., Яковлев А. С., Алексеев Д. А. Раковинные корненожки (Testacida) почв // Почвенные простейшие . Ленинград: Наука, 1980. С. 108—142.
Цыганов А. Н., Комаров А. А., Мазей Н. Г., Борисова Т. В. Динамика видовой структуры сообщества раковинных амеб в ходе сукцессии “водоем–болото” в голоцене на примере болота Мочуля (Калужская область, Россия) // Зоологический журнал. 2020. T. 99. № 5. С. 586—598.
Рахлеева А. А., Семенова Т. А., Стриганова Б. Р., Терехова В. А. Динамика зоомикробных комплексов при разложении растительного опада в ельниках южной тайги // Почвоведение. 2011. № 1. С. 44—55.
Кац Н. Я. Болота земного шара. Москва: Наука, 1971. 296 с.
Корганова Г. А. Почвенные раковинные амёбы (Protozoa: Testacea): фауна, экология, принципы организации сообществ: диссертация доктора биол. наук. Москва, 1997. 340 с.
Коросов А. В. Специальные методы биометрии. Петрозаводск, 2007. 364 с.
Красильников П. В., Платонова Е. А. Почвы заповедной территории Ботанического сада ПетрГУ // Hortus Bot. 2001. № 1. С. 34—41.
Кулюкина Е. В. Влияние антропогенной нагрузки на численный и видовой состав раковинных амеб в почвах г. Томска // Научно-исследовательские публикации: природа, экология и народное хозяйство. 2015. Т. 1. № 7 (27). С. 59—64.
Кулюкина Е. В., Карташев А. Г., Денисова Т. В. Пространственное распределение раковинных амеб в ризосфере сосны и ели // Вестник Российского университета дружбы народов. Экология и безопасность. 2016. № 4. С. 18—32.
Мазей Ю. А., Бубнова О. А., Чернышев В. А. Структура сообщества раковинных амеб (Testacealobosea; Testaceafilosea; Amphitremidae) в Чибирлейском моховом болоте (среднее Поволжье) // Известия Самарского научного центра РАН. 2009. Т. 11. № 1. С. 72—77.
Мазей Ю. А., Ембулаева Е. А. Изменение сообществ почвообитающих раковинных амеб вдоль лесостепного градиента в Среднем Поволжье // Аридные экосистемы. 2009. Т. 15. № 37. С. 13—23.
Мазей Ю. А., Кабанов А. Н. Раковинные амебы в осоковосфагновом заболоченном лесу на севере Карелии // Известия Пензенского государственного педагогического университета им. В. Г. Белинского. 2008. № 14. С. 101—104.
Мазей Ю. А., Комаров А. А. Видовой состав и распределение раковинных амеб в соответствии с микромозаичной организацией основных типов темнохвойных лесов в верховьях р. Печоры // Труды Печоро-Илычского заповедника. 2015. № 17. С. 120—125.
Мазей Ю. А., Цыганов А. Н. Пресноводные раковинные амебы. Москва: Товарищество научных изданий КМК, 2006. 300 с.
Мазей Ю. А., Цыганов А. Н., Бубнова О. А. Видовой состав, распределение и структура сообщества раковинных амеб мохового болота в Среднем Поволжье // Зоологический журнал. 2007. Т. 86. Вып. 10. С. 1155—1167.
Мазей Ю. А, Цыганов А. Н., Бубнова О. А. Структура сообщества раковинных амеб в заболоченных биотопах южной тайги Европейской части России // Успехи современной биологии. 2009. Т. 129. № 1. С. 67—77.
Песенко Ю. А. Принципы и методы количественного анализа в фаунистических исследованиях. Москва: Наука, 1982. 286 с.
Рахлеева А. А., Корганова Г. А. К вопросу об оценке численности и видового разнообразия раковинных амеб (Rhizopoda, Testacea) в таежных почвах // Зоологический журнал. 2005. Т. 84. № 12. С. 1427—1436.
Смолянинова С. Э., Гренадерова А. В. Сообщества раковинных амеб в подстилке сосняков Юксеевского лесничества после воздействия пожара // Вестник Красноярского государственного аграрного университета. 2018. №5. С. 307—313.
Бабешко К. В., Цыганов А. Н., Новенко Е. Ю. и др. Современные и ископаемые сообщества раковинных амеб в болотных экосистемах национального парка «Орловское полесье» // Ученые записки Орловского государственного университета. 2015. № 3 (59). С. 302—310.
Трулова А. С. , Мазей Ю. А. Сезонная динамика структуры сообщества раковинных амеб в Среднем Поволжье // Известия ПГПУ им. В. Г. Белинского. 2012. № 29. С. 397—404.
Bobrov A. A., Charman D. J., Warner B. G. Ecology of Testate Amoebae from Oligotrophic Peatlands: Specific Features of Polytypic and Polymorphic Species // Biology Bulletin of the Russian Academy of Sciences. 2002. Vol. 29. P. 605—617.
Bo-Ping Han, Tian Wang, Lei Xu, Qiu Qi Lin, Zhang Jinyu, Henri J. Dumont Carnivorous planktonic Difflugia (Protista, Amoebina Testacea) and their predators // European Journal of Protistology. 2011. Vol. 47. P. 214—223.
Chambers F. M., Beilman D. W. Methods for determining peat humification and for quantifying peat bulk density, organic matter and carbon content for palaeostudies of climate and peatland // Mires and Peat. 2010. Vol. 7. P. 1—10.
Charman D. J., D. Hendon, W. A. Woodland The identification of testate amoebae (Protozoa: Rhizopoda) in peats // Quaternary Research Association. London, 2000. 200 p.
Robert L. Ellison Paleolimnological analysis of Ullswater using testate amoebae // Journal of Paleolimnology. 1995. Vol. 13. P. 51—63.
