

В связи с активным привлечением в зелёное строительство видов природной флоры возникла необходимость в анализе, выявляющем характер взаимоотношений между растительными организмами и факторами окружающей среды их новых местопроизрастаний.
В частности, особенности репродуктивной биологии вида целесообразно определять, учитывая влияние освещённости, так как, в зависимости от условий светового режима и качественного состава света, у растений меняется биосинтез функционально-активных продуктов, что приводит к изменению морфогенеза (Куперман, 1978; Ничипорович и др., 1985). Наличие и регулярность плодоношения, качественные и количественные показатели семян свидетельствуют о приспособленности и адаптации вида к данным условиям произрастания.
Виды рода Rhodiola L. (Родиола) семейства Crassulaceae DC. растут в горных системах Центральной Азии, Восточной и Западной Европы, на Памире, Тянь-Шане, Алтае, в Саянах, реже – в Северной Америке на каменистых почвах, по скалам, на лесных лугах, альпийских лужайках (Флора Казахстана, 1961; Флора Западной Сибири, 1964; Краснов и др., 1979). Засухостойчивы, морозостойки, нетребовательны к плодородию почвы и декоративны в течение длительного периода (Привалко, 2010), поэтому заслуживают более широкого использования в фитодизайне.
R. kirilowii (Regel) Maxim. в условиях культуры в Донецком ботаническом саду (ДБС) ранее успешно прошла интродукционное испытание и получила высокую оценку декоративности при выращивании как на хорошо освещённых, так и на затенённых участках (Привалко, 2010).
Цель работы – изучить онтогенез R. kirilowii (Regel) Maxim. первых 7-ми лет жизни и выявить особенности её репродуктивной биологии в условиях разной освещённости в степной зоне. Для этого определяли периоды прохождения растениями возрастных состояний, их морфометрические параметры и время вступления в период декоративности, проанализировали наличие и регулярность плодоношения, качественные и количественные показатели семян, а также укореняемость черенков в условиях защищённого грунта в песке и почвосмеси и сразу в открытом грунте на освещённых и затенённых участках.
R. kirilowii – вегетативно неподвижный стержнекорневой короткокорневищный многолетник; гемикриптофит; мало- или умеренно требовательный к плодородию почвы; энтомофильный барохорный эуксерофит; сциогелиофит, петрофит, хасмофит (Raunkiaer C., 1907; Серебряков И. Г., 1962; Зиман С. Н., 1976; Бельгардт А. Л., 1980; Тарасов В. В., 2005). Экологически приурочена к трещинам скал, склонам, перевалам Средней Азии (Тянь-Шань, Памиро-Алай, Тибет) (Борисова А. Г., 1970).
Мобилизацию объекта исследования провели семенами, полученными в 2003 году из Мюнхенского ботанического сада (Германия). Видовое название указано с учетом современных номенклатурных сводок (The Plant List, 2013).
ДБС расположен в Донецком округе Приазовско-Черноморской степной и Причерноморской степной провинций Европейско-Азиатской степной зоны Голарктического Доминиона (Геоботанічне районування Української РСР, 1977). Для района характерна годовая суммарная солнечная радиация 1200–1400 кВт/м2. Среднегодовое количество осадков 450–520 мм, из них за вегетационный период выпадает 260–310 мм. Наибольшее их количество приходится на первую половину лета, наименьшее – на февраль. Количество засушливых дней – 38–59. Засухи и суховеи отмечаются преимущественно во второй половине лета, а в отдельные годы и весной. Средняя годовая температура воздуха +6,0–7,8o С. Продолжительность периода со среднесуточными температурами выше +10o С составляет 165–170 дней. Сумма температур за этот период равна 2900–3100o С. Последние весенние заморозки прекращаются в среднем 21–28 апреля, а первые осенние начинаются 6–12 октября. Зимы чаще всего малоснежные, с неустойчивым снеговым покровом и частыми оттепелями. Иногда наблюдаются резкие перепады температур, приводящие к образованию изморозей и гололеда (Бабиченко и др., 1984).
Онтогенетическое развитие R. kirilowii изучали в течение 7 лет в соответствии с классификацией Т. А. Работнова (Работнов, 1964), дополненной А. А. Урановым (Уранов, 1960) с использованием общепринятых методик (Игнатьева, 1964; Смирнова и др., 1976). По мере перехода растений в очередное возрастное состояние 2–3 особи выкапывали и определяли морфометрические параметры. Для определения особенностей образования морфологических структур проводили описание морфогенеза органов растений, выращенных из семян местной репродукции. Терминологию периодов онтогенеза и возрастных состояний особей приводили по Л. И. Воронцовой (Воронцова и др., 1976). Семена в начале января проращивали на свету в чашках Петри, сеянцы пикировали в садовую почву с площадью питания 1x1 см и выращивали в условиях теплицы, в конце первого года вегетации 100 модельных особей высадили в открытый грунт на незатенённые участки. Растения периодически выкапывали (по 3–5 штук одновременно) и отмечали изменения, произошедшие в строении надземной и подземной частей со времени предыдущего наблюдения.
Для определения у R. kirilowii, произрастающей в разных световых режимах, семенной продуктивности, лабораторной всхожести, массы 1000 семян и количества семян в 1 мг, на территории ДБС заложили 2 варианта экспериментальных участков – освещенные и затенённые – в трех повторностях. На все участки высадили по 30 шт. предварительно укорененных черенков. Растения выращивали без дополнительного полива, в условиях естественной длины дня и интенсивности освещения. Степень освещенности участков измеряли люксметром Ю–116 ежедневно в 815, 1200 и 1545.
Семенную продуктивность, лабораторную всхожесть, массу 1000 семян и количество семян в 1 мг изучали по методике И. В. Вайнагий (Вайнагий, 1974). Также использовали методические указания по семеноводству интродуцентов (Методические указания по семеноводству интродуцентов, 1980) и методы изучения семенного размножения травянистых растений в сообществах (Работнов, 1960). При определении реальной семенной продуктивности учитывали количество генеративных побегов на особь, плодолистиков на одном растении и семян, формирующихся в одном плодолистике. Для определения лабораторной всхожести семян их проращивание проводили в чашках Петри на влажной фильтровальной бумаге в трехкратной повторности по 50 штук для каждого варианта опыта. Подсчет проросших семян вели через день.
Для определения оптимальных условий вегетативного размножения в июне – июле проводили одновременное укоренение черенков в условиях защищенного и открытого грунта на затенённых и незатенённых участках. Все цветоносы и нижние листья удалили. Для черенкования в условиях защищённого грунта использовали два вида субстрата: песок и почвосмесь (садовая почва и перегной, 1:1). Полив проводили ежедневно водопроводной водой. В открытом грунте черенки укореняли непосредственно в почве без мульчирования и использования укрывных материалов, с поливом участков 2–3 раза в неделю. Укоренение длилось в течение месяца. Затем определяли укореняемость черенков и размеры образованных корневых систем (максимальную длину корней в корневой системе и максимальное ее распространение в ширину).
В условиях ДБС за 7 лет все особи R. kirilowii прошли этапы развития от латентного периода до генеративного и достигли зрелого генеративного возраста. Отмечено ежегодное плодоношение генеративных особей, при этом самосев был единичным и нерегулярным. К концу первого года жизни сеянцы находились в имматурном состоянии, за второй год прошли через виргинильное, за третий и четвертый – молодое генеративное, с пятого по седьмой находились в зрелом генеративном. То есть с третьего по седьмой годы жизни были декоративны и пригодны для использования в декоративных фитонасаждениях.
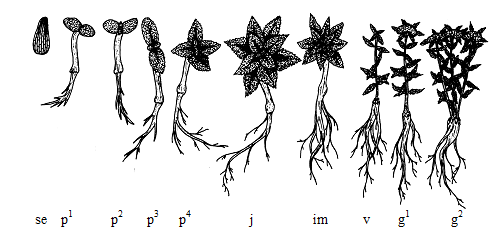
Рис. 1. Схема стадий онтогенеза Rhodiola kirilowii (Regel) Maxim. первых 7 лет жизни.
Латентный период: se – семя. Прегенеративный период: p1 – проростки – на 14 день, p2 – на 50 день, p3 – на 70 день, p4 – на 90 день; j – ювенильное состояние, im – имматурное состояние, v – виргинильное состояние растений. Генеративный период: g1 – молодая генеративная особь, g2 – зрелая генеративная особь.
Fig. 1. Diagram of ontogeny stages of Rhodiola kirilowii (Regel) Maxim. in the first 7 years of life.
Latent period: se - seed. Pregenerative period: p1 - seedlings (14th day), p2 (50th day), p3 (70th day), p4 (90th day); j - juvenile condition, im - immature status, v - virginal state of plants. Generative period: g1 - young generative individual, g2 - mature generative individual.
Латентный период
Семена яйцевидные, длиной около 2 мм, коричневые, с темно-коричневыми продольными бороздками (рис. 1). Всхожесть семян в лабораторных условиях составляет 21–32 %. Массовое прорастание семян происходит на 7–9 день. Прегенеративный период.
Проростки. Семядоли вначале светло-зеленые, у двухнедельных проростков – ярко-зеленые, яйцевидные, размером 1,5×1,0 мм, мясистые, голые, цельнокрайние, сохраняются 45–55 дней. Гипокотиль длиной 0,6–0,8 мм. Развитие боковых корней первого порядка отмечено на 10–14 день после прорастания семян, второго порядка – на 65–75 день, а третьего – на 80–90 день. Конус нарастания (почечка) появляется на 30–35 день. Первая пара настоящих листьев начинает разворачиваться на 50–60 день и достигает своих максимальных размеров на 65–75 день. Параллельно (на 60–70 день) развивается вторая пара листьев. Листорасположение супротивное, междоузлия короткие. Листья мясистые, сидячие, в первой паре – округло-яйцевидной формы, почти цельнокрайние, на вершине слегка заостренные. Следующие одна–две пары листьев яйцевидные, слабозубчатые, на вершине заостренные. Начиная с четвертой-пятой пары листьев форма листовой пластинки становится линейно-ланцетной, край – зубчатым, вершина – острой.
Ювенильное состояние. На 80–90 день развития растения достигли ювенильного состояния и характеризовались потерей семядолей, появлением боковых корней третьего порядка и наличием настоящих листьев нехарактерной для взрослых растений формы.
Имматурное состояние. Отмечено на 125–140 день после появления всходов: развились боковые корни четвертого порядка, появились листья характерной для взрослых растений формы, но отличающиеся по размерам и расположенные в коротких междоузлиях. На 270 день растения были 4–5 см высотой, имели боковые корни третьего – четвертого порядков и по 6–7 пар настоящих листьев (из них 2–4 пары типичных для данного вида).
Виргинильное состояние. Наступило и продолжалось в течение второго года жизни. Появились основные характерные для взрослого растения черты (побеги, листья, корни, жизненная форма). Цветение не отмечено.
Генеративный период
Молодые генеративные особи. За третий и четвертый годы жизни произошло окончательное становление жизненной формы, цветение и плодоношение стало регулярным. Цветение начинается в конце мая – первой половине июня и длится до 40 дней. Корневая система хорошо развита и имеет вид утолщенного вертикального стержня с системой боковых корней. В верхней части главного корня имеются чешуевидные пленчатые треугольные острые листья. Растения достигают 38–45 см в высоту и образуют по 2–5 вегетативных однолетних побегов, заканчивающихся соцветием. В соцветиях имеется в среднем по 20–40 цветков. Длина листовой пластинки достигает 5,8–6,1 см, ширина – 1,1–1,7 см. Плодоношение ежегодное.
Зрелые генеративные особи. На пятый год жизни отмечен максимальный прирост надземной массы, обильное цветение и плодоношение. Главный корень увеличился в диаметре. Высота растений составила 30–50 см, количество вегетативных (= генеративных) побегов – 2–6, в соцветиях – по 25–50 цветков. Длина листовой пластинки – 6,0–6,5, ширина – 1,3–1,8 см. В течение шестого и седьмого годов жизни морфометрические показатели существенно не изменились. Плодоношение ежегодное.
Для определения влияния светового режима местопроизрастания R. kirilowii на особенности её размножения семенным способом изучена взаимосвязь между условиями освещенности местопроизрастания и лабораторной всхожестью, массой 1000 семян и количеством семян в 1 г, а также реальной семенной продуктивностью.
На незатенённых экспериментальных участках минимальная среднемесячная освещённость отмечена в январе (≈17000 люкс), максимальная – в июле (≈36000 люкс), максимальная за год – ≈23000 люкс, минимальная за год – ≈20000 люкс; на затенённых – соответственно ≈8000 люкс, ≈19000 люкс, ≈10000 люкс и ≈11000 люкс.
Таблица 1. Репродуктивные показатели Rhodiola kirilowii (Regel) Maxim. в условиях разной освещенности местопроизрастаний
Table 1. Reproductive performance of Rhodiola kirilowii (Regel) Maxim. under different habitat illumination
| Репродуктивные показатели | Высокая освещённость | Затенение | t-критерий Стьюдента |
| Масса 1000 семян, мг, M±m | 8,00±0,01 | 11,53±0,08 | 44,13*** |
| Количество семян в 1 г, шт., M±m | 12376±0,03 | 8680±0,04 | 12,83*** |
| Реальная семенная продуктивность | 2766,10±6,32 | 1354,00±65,62 | 15,33*** |
Примечания: M±m – среднее арифметическое ± ошибка среднего;
различия достоверны при Р≥0,999 (***).
Notes: M ± m is an arithmetic mean ± error of the mean;
the differences were significant at P≥0,999 (***).
Выявлено, что у семян, собранных на затенённых участках, лабораторная всхожесть в 2 раза ниже чем на незатенённых (21,56 % и 43,50 % соответственно). С уменьшением уровня освещённости местопроизрастания у R. kirilowii достоверно снижается и реальная семенная продуктивность (табл. 1). При этом в затенении масса 1000 семян достоверно больше, а их количество в 1 г меньше. Поэтому для семенного размножения R. kirilowii маточники рекомендуется выращивать на участках с высокой освещённостью.
Поскольку при выращивании из семян молодые растения R. kirilowii декоративны только с третьего года жизни, для зелёного строительства семенное размножение данного вида экономически менее целесообразно, чем размножение методом черенкования.
При укоренении черенков R. kirilowii в условиях защищённого грунта в песке и почвосмеси и сразу в открытом грунте на освещённых и затенённых участках нами отмечена 100 % укореняемость черенков во всех вариантах эксперимента.
При анализе размеров образованной корневой системы выявлено, что способ укоренения не влияет на её длину, а максимального распространения в ширину корневая система достигает в песке в условиях защищённого грунта (табл. 2).
Таблица 2. Размеры корневой системы, образованной при укоренении черенков Rhodiola kirilowii (Regel) Maxim. разными способами
Table 2. The size of the root system formed in the process of rooting Rhodiola kirilowii (Regel) Maxim. cuttings by different means
| Размеры корневой системы, см, M±m | В песке | В почвосмеси | В открытом грунте | |
| высокая освещённость | затенение | |||
| Максимальная длина корневой системы | 7,13±0,35 | 7,07±0,65 | 7,01±0,82 | 7,02±0,93 |
| Максимальное распространение корневой системы в ширину | 5,65±0,37 | 4,10±0,40 | 3,96±0,74 | 4,02±0,61 |
Примечания: M±m – среднее арифметическое ± ошибка среднего.
Notes: M ± m is an arithmetic mean ± error of the mean.
Следовательно, данный вид целесообразнее укоренять сразу на постоянном месте в цветочной композиции, независимо от освещённости будущего местопроизрастания, желательно в почве с добавлением песка.
В течение 7 лет в условиях Донецкого ботанического сада R. kirilowii проходит этапы развития от латентного периода до зрелого генеративного возраста. С третьего по седьмой годы жизни растения декоративны. Плодоношение генеративных особей ежегодное, при этом самосев единичный и нерегулярный. Для семенного размножения R. kirilowii маточники рекомендуется выращивать на участках с высокой освещённостью. При вегетативном размножении данный вид целесообразнее укоренять сразу на постоянном месте в цветочной композиции, независимо от освещённости будущего местопроизрастания, желательно в почве с добавлением песка.
References
Babitchenko V. N., Barabash M. B., Logvinov K. T. i dr. Priroda Ukrainskoj SSR: Klimat. Nature of the Ukrainian SSR: The climate Kiev: Nauk. dumka, 1984. 232 s.
Belgardt A. L. K voprosu ob ekologitcheskom analize i strukture lesnykh fitotsenozov v stepi // Voprosy biologitcheskoj diagnostiki lesnykh biogeotsenozov Primorya. On environmental analysis and structure of forest communities in the steppe // Issues of biological diagnostics of Primorye forest ecosystems Dnepropetrovsk, 1980. S. 12—43.
Borisova A. G. Konspekt sistemy sem. Crassulaceae DC. flory SSSR (dobavleniya i izmeneniya) // Novosti sistematiki vysshikh rastenij. Synopsis of the system of Crassulaceae DC. family of USSR flora (add. and edit.) // News of systematics of higher plants L., 1969. T. 6. S. 112—121.
Vajnagij I. V. O metodike izutcheniya semennoj produktivnosti rastenij // Botan. zhurn. On the method of studying the seed production of plants // Botan. Journ. 1974. T. 59. № 6. S. 826—830.
Vorontsova L. I., Gattsuk L. E., Egorova V. N. i dr. Tsenopopulyatsii rastenij (osnovnye ponyatiya i struktura). Plant cenopopulations (basic concepts and structure) M.: Nauka, 1976. 214 s.
Geobotanіtchne rajonuvannya Ukraїnskoї RSR. Geobotanical zoning of the Ukrainian SSR K.: Nauk. dumka, 1977. 302 s.
Ziman S. N. Zhiznennye formy i biologiya stepnykh rastenij Donbassa. Life forms and biology of Donbass steppe plants Kiev: Nauk. dumka, 1976. 190 s.
Ignateva I. P. Metodika izutcheniya morfogeneza vegetativnykh organov travyanistykh polikarpikov // Dokl. TSKhA. Methods of study of morphogenesis of herbaceous vegetative organs polycarpic // Reports of TSKHA 1964. № 98. S. 47—57.
Krasnov E. A., Saratikov A. S., Surov Yu. P. Rasteniya semejstva Tolstyankovykh. The plants of the family Crassulaceae Tomsk, 1979. 207 s.
Kuperman F. M. Issledovanie zakonomernostej morfogeneza rastenij metodom vyratshivaniya ikh v usloviyakh raznykh svetovykh rezhimov // Svet i morfogenez rastenij / Pod red. F. M. Kupermana, E. I. Rzhanovskoj. Research of morphogenesis regularities of plants by growing them in different light conditions // Light and morphogenesis of plants / Ed. F. M. Kuperman, E. I. Rzhanovskoy M.: Izd-vo Mosk. un-ta, 1978. S. 8—43.
Metoditcheskie ukazaniya po semenovodstvu introdutsentov / Otv. red. N. V. Tsitsin. Guidelines for seed breeding of introduced plants / Ed. N. V. Tsitsin M., 1980. 64 s.
Nitchiporovitch A. A., Kefeli V. I., Aknazarov O. A. Dejstvie svetovykh faktorov vysokogorij Pamira na zhiznedeyatelnost rastenij. The effect of light factors in the high mountains of the Pamirs on plant life Dushanbe: Donish, 1985. 216 s.
Privalko L. V. Dekorativnye tolstyankovye dlya ispolzovaniya v landshaftnom fitodizajne: rekomendatsii / Pod obtsh. red. A. Z. Glukhova. Decorative Crassulaceae plants for use in landscaping and phytodesign: Recommendations / Ed. A. Z. Glukhov Donetsk, 2010. 20 s.
Rabotnov T. A. Metody izutcheniya semennogo razmnozheniya travyanistykh rastenij v soobtshestvakh // Polevaya geobotanika. Methods of study of herbaceous plants seed breeding in communities // Field geobotany M., L.: Nauka, 1960. T. 2. S. 20—40.
Rabotnov T. A. Opredelenie vozrastnogo sostava populyatsij vidov v soobtshestve // Polevaya geobotanika. Determination of the age composition of species populations in the community // Field geobotany M., L.: Nauka, 1964. T. 3. S. 132—208.
Rabotnov T. A. Zhiznennyj tsikl mnogoletnikh travyanistykh rastenij v lugovykh tsenozakh // Tr. BIN AN SSSR. Life cycle of perennial herbaceous plants in the grassland cenoses // Proceedings of BIN USSR Academy of Sciences 1965. Ser. 3. № 6. S. 7—204.
Serebryakov I. G. Ekologitcheskaya morfologiya rastenij. Zhiznennye formy pokrytosemennykh i khvojnykh. Ecological plant morphology. Life forms of angiosperms and conifers M.: Vyssh. shk., 1962. 378 s.
Smirnova O. V., Zaugolnova L. B., Toronova N. A. Kriterii vydeleniya vozrastnykh sostoyanij i osobennosti khoda ontogeneza u rastenij razlitchnykh biomorf // Tsenopopulyatsiya rastenij. Criteria for determination of age-stages and specific features of ontogeny in plants of different biomorphs // Cenotic populations of plants M.: Nauka, 1976. S. 14—43.
Tarasov V. V. Flora Dnіpropetrovskoї ta Zaporіzkoї oblastej. Sudinnі roslini. Bіologo-ekologіtchna kharakteristika vidіv. Flora of Dnepropetrovsk and Zaporozhye regions. Vascular plants. Biology and environmental characteristics of the species D.: Vid-vo DNU, 2005. 276 s.
Uranov A. A. Zhiznennye sostoyaniya vida v rastitelnom soobtshestve // Byull. MOIP. Ser. Biol. Vital stages of a species in plant community // Bull. Moscow Society of Naturalists. Ser. Biol. 1960. Vyp. 3. S. 77—92.
Flora Zapadnoj Sibiri. Flora of West Siberia Tomsk, 1964. T. 12. 327 s.
Flora Kazakhstana: v 9 t. Flora of Kazakhstan: 9 vols. Alma-Ata, 1961. T. 4. 344 s.
Raunkiaer C. Planterigets Livsformer of deres Betydning for Geografien. Kobenhavn: Nordiskforland, 1907. 132 p.
The Plant List, 2013. Version 2; URL: http://www.theplantlist.org/ (data obratsheniya 20.02.2017).
