

Введение
В связи с активным привлечением в зелёное строительство видов природной флоры возникла необходимость в анализе, выявляющем характер взаимоотношений между растительными организмами и факторами окружающей среды их новых местопроизрастаний.
В частности, особенности репродуктивной биологии вида целесообразно определять, учитывая влияние освещённости, так как, в зависимости от условий светового режима и качественного состава света, у растений меняется биосинтез функционально-активных продуктов, что приводит к изменению морфогенеза (Куперман, 1978; Ничипорович и др., 1985). Наличие и регулярность плодоношения, качественные и количественные показатели семян свидетельствуют о приспособленности и адаптации вида к данным условиям произрастания.
Виды рода Rhodiola L. (Родиола) семейства Crassulaceae DC. растут в горных системах Центральной Азии, Восточной и Западной Европы, на Памире, Тянь-Шане, Алтае, в Саянах, реже – в Северной Америке на каменистых почвах, по скалам, на лесных лугах, альпийских лужайках (Флора Казахстана, 1961; Флора Западной Сибири, 1964; Краснов и др., 1979). Засухостойчивы, морозостойки, нетребовательны к плодородию почвы и декоративны в течение длительного периода (Привалко, 2010), поэтому заслуживают более широкого использования в фитодизайне.
R. kirilowii (Regel) Maxim. в условиях культуры в Донецком ботаническом саду (ДБС) ранее успешно прошла интродукционное испытание и получила высокую оценку декоративности при выращивании как на хорошо освещённых, так и на затенённых участках (Привалко, 2010).
Цель работы – изучить онтогенез R. kirilowii (Regel) Maxim. первых 7-ми лет жизни и выявить особенности её репродуктивной биологии в условиях разной освещённости в степной зоне. Для этого определяли периоды прохождения растениями возрастных состояний, их морфометрические параметры и время вступления в период декоративности, проанализировали наличие и регулярность плодоношения, качественные и количественные показатели семян, а также укореняемость черенков в условиях защищённого грунта в песке и почвосмеси и сразу в открытом грунте на освещённых и затенённых участках.
Объекты и методы исследований
R. kirilowii – вегетативно неподвижный стержнекорневой короткокорневищный многолетник; гемикриптофит; мало- или умеренно требовательный к плодородию почвы; энтомофильный барохорный эуксерофит; сциогелиофит, петрофит, хасмофит (Raunkiaer C., 1907; Серебряков И. Г., 1962; Зиман С. Н., 1976; Бельгардт А. Л., 1980; Тарасов В. В., 2005). Экологически приурочена к трещинам скал, склонам, перевалам Средней Азии (Тянь-Шань, Памиро-Алай, Тибет) (Борисова А. Г., 1970).
Мобилизацию объекта исследования провели семенами, полученными в 2003 году из Мюнхенского ботанического сада (Германия). Видовое название указано с учетом современных номенклатурных сводок (The Plant List, 2013).
ДБС расположен в Донецком округе Приазовско-Черноморской степной и Причерноморской степной провинций Европейско-Азиатской степной зоны Голарктического Доминиона (Геоботанічне районування Української РСР, 1977). Для района характерна годовая суммарная солнечная радиация 1200–1400 кВт/м2. Среднегодовое количество осадков 450–520 мм, из них за вегетационный период выпадает 260–310 мм. Наибольшее их количество приходится на первую половину лета, наименьшее – на февраль. Количество засушливых дней – 38–59. Засухи и суховеи отмечаются преимущественно во второй половине лета, а в отдельные годы и весной. Средняя годовая температура воздуха +6,0–7,8o С. Продолжительность периода со среднесуточными температурами выше +10o С составляет 165–170 дней. Сумма температур за этот период равна 2900–3100o С. Последние весенние заморозки прекращаются в среднем 21–28 апреля, а первые осенние начинаются 6–12 октября. Зимы чаще всего малоснежные, с неустойчивым снеговым покровом и частыми оттепелями. Иногда наблюдаются резкие перепады температур, приводящие к образованию изморозей и гололеда (Бабиченко и др., 1984).
Онтогенетическое развитие R. kirilowii изучали в течение 7 лет в соответствии с классификацией Т. А. Работнова (Работнов, 1964), дополненной А. А. Урановым (Уранов, 1960) с использованием общепринятых методик (Игнатьева, 1964; Смирнова и др., 1976). По мере перехода растений в очередное возрастное состояние 2–3 особи выкапывали и определяли морфометрические параметры. Для определения особенностей образования морфологических структур проводили описание морфогенеза органов растений, выращенных из семян местной репродукции. Терминологию периодов онтогенеза и возрастных состояний особей приводили по Л. И. Воронцовой (Воронцова и др., 1976). Семена в начале января проращивали на свету в чашках Петри, сеянцы пикировали в садовую почву с площадью питания 1x1 см и выращивали в условиях теплицы, в конце первого года вегетации 100 модельных особей высадили в открытый грунт на незатенённые участки. Растения периодически выкапывали (по 3–5 штук одновременно) и отмечали изменения, произошедшие в строении надземной и подземной частей со времени предыдущего наблюдения.
Для определения у R. kirilowii, произрастающей в разных световых режимах, семенной продуктивности, лабораторной всхожести, массы 1000 семян и количества семян в 1 мг, на территории ДБС заложили 2 варианта экспериментальных участков – освещенные и затенённые – в трех повторностях. На все участки высадили по 30 шт. предварительно укорененных черенков. Растения выращивали без дополнительного полива, в условиях естественной длины дня и интенсивности освещения. Степень освещенности участков измеряли люксметром Ю–116 ежедневно в 815, 1200 и 1545.
Семенную продуктивность, лабораторную всхожесть, массу 1000 семян и количество семян в 1 мг изучали по методике И. В. Вайнагий (Вайнагий, 1974). Также использовали методические указания по семеноводству интродуцентов (Методические указания по семеноводству интродуцентов, 1980) и методы изучения семенного размножения травянистых растений в сообществах (Работнов, 1960). При определении реальной семенной продуктивности учитывали количество генеративных побегов на особь, плодолистиков на одном растении и семян, формирующихся в одном плодолистике. Для определения лабораторной всхожести семян их проращивание проводили в чашках Петри на влажной фильтровальной бумаге в трехкратной повторности по 50 штук для каждого варианта опыта. Подсчет проросших семян вели через день.
Для определения оптимальных условий вегетативного размножения в июне – июле проводили одновременное укоренение черенков в условиях защищенного и открытого грунта на затенённых и незатенённых участках. Все цветоносы и нижние листья удалили. Для черенкования в условиях защищённого грунта использовали два вида субстрата: песок и почвосмесь (садовая почва и перегной, 1:1). Полив проводили ежедневно водопроводной водой. В открытом грунте черенки укореняли непосредственно в почве без мульчирования и использования укрывных материалов, с поливом участков 2–3 раза в неделю. Укоренение длилось в течение месяца. Затем определяли укореняемость черенков и размеры образованных корневых систем (максимальную длину корней в корневой системе и максимальное ее распространение в ширину).
Результаты и обсуждение
В условиях ДБС за 7 лет все особи R. kirilowii прошли этапы развития от латентного периода до генеративного и достигли зрелого генеративного возраста. Отмечено ежегодное плодоношение генеративных особей, при этом самосев был единичным и нерегулярным. К концу первого года жизни сеянцы находились в имматурном состоянии, за второй год прошли через виргинильное, за третий и четвертый – молодое генеративное, с пятого по седьмой находились в зрелом генеративном. То есть с третьего по седьмой годы жизни были декоративны и пригодны для использования в декоративных фитонасаждениях.
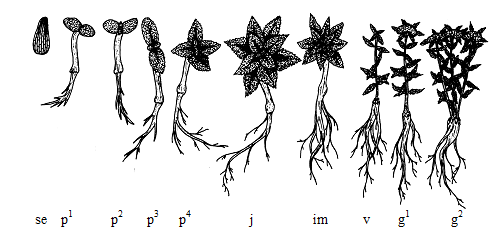
Рис. 1. Схема стадий онтогенеза Rhodiola kirilowii (Regel) Maxim. первых 7 лет жизни.
Латентный период: se – семя. Прегенеративный период: p1 – проростки – на 14 день, p2 – на 50 день, p3 – на 70 день, p4 – на 90 день; j – ювенильное состояние, im – имматурное состояние, v – виргинильное состояние растений. Генеративный период: g1 – молодая генеративная особь, g2 – зрелая генеративная особь.
Латентный период
Семена яйцевидные, длиной около 2 мм, коричневые, с темно-коричневыми продольными бороздками (рис. 1). Всхожесть семян в лабораторных условиях составляет 21–32 %. Массовое прорастание семян происходит на 7–9 день. Прегенеративный период.
Проростки. Семядоли вначале светло-зеленые, у двухнедельных проростков – ярко-зеленые, яйцевидные, размером 1,5×1,0 мм, мясистые, голые, цельнокрайние, сохраняются 45–55 дней. Гипокотиль длиной 0,6–0,8 мм. Развитие боковых корней первого порядка отмечено на 10–14 день после прорастания семян, второго порядка – на 65–75 день, а третьего – на 80–90 день. Конус нарастания (почечка) появляется на 30–35 день. Первая пара настоящих листьев начинает разворачиваться на 50–60 день и достигает своих максимальных размеров на 65–75 день. Параллельно (на 60–70 день) развивается вторая пара листьев. Листорасположение супротивное, междоузлия короткие. Листья мясистые, сидячие, в первой паре – округло-яйцевидной формы, почти цельнокрайние, на вершине слегка заостренные. Следующие одна–две пары листьев яйцевидные, слабозубчатые, на вершине заостренные. Начиная с четвертой-пятой пары листьев форма листовой пластинки становится линейно-ланцетной, край – зубчатым, вершина – острой.
Ювенильное состояние. На 80–90 день развития растения достигли ювенильного состояния и характеризовались потерей семядолей, появлением боковых корней третьего порядка и наличием настоящих листьев нехарактерной для взрослых растений формы.
Имматурное состояние. Отмечено на 125–140 день после появления всходов: развились боковые корни четвертого порядка, появились листья характерной для взрослых растений формы, но отличающиеся по размерам и расположенные в коротких междоузлиях. На 270 день растения были 4–5 см высотой, имели боковые корни третьего – четвертого порядков и по 6–7 пар настоящих листьев (из них 2–4 пары типичных для данного вида).
Виргинильное состояние. Наступило и продолжалось в течение второго года жизни. Появились основные характерные для взрослого растения черты (побеги, листья, корни, жизненная форма). Цветение не отмечено.
Генеративный период
Молодые генеративные особи. За третий и четвертый годы жизни произошло окончательное становление жизненной формы, цветение и плодоношение стало регулярным. Цветение начинается в конце мая – первой половине июня и длится до 40 дней. Корневая система хорошо развита и имеет вид утолщенного вертикального стержня с системой боковых корней. В верхней части главного корня имеются чешуевидные пленчатые треугольные острые листья. Растения достигают 38–45 см в высоту и образуют по 2–5 вегетативных однолетних побегов, заканчивающихся соцветием. В соцветиях имеется в среднем по 20–40 цветков. Длина листовой пластинки достигает 5,8–6,1 см, ширина – 1,1–1,7 см. Плодоношение ежегодное.
Зрелые генеративные особи. На пятый год жизни отмечен максимальный прирост надземной массы, обильное цветение и плодоношение. Главный корень увеличился в диаметре. Высота растений составила 30–50 см, количество вегетативных (= генеративных) побегов – 2–6, в соцветиях – по 25–50 цветков. Длина листовой пластинки – 6,0–6,5, ширина – 1,3–1,8 см. В течение шестого и седьмого годов жизни морфометрические показатели существенно не изменились. Плодоношение ежегодное.
Для определения влияния светового режима местопроизрастания R. kirilowii на особенности её размножения семенным способом изучена взаимосвязь между условиями освещенности местопроизрастания и лабораторной всхожестью, массой 1000 семян и количеством семян в 1 г, а также реальной семенной продуктивностью.
На незатенённых экспериментальных участках минимальная среднемесячная освещённость отмечена в январе (≈17000 люкс), максимальная – в июле (≈36000 люкс), максимальная за год – ≈23000 люкс, минимальная за год – ≈20000 люкс; на затенённых – соответственно ≈8000 люкс, ≈19000 люкс, ≈10000 люкс и ≈11000 люкс.
Таблица 1. Репродуктивные показатели Rhodiola kirilowii (Regel) Maxim. в условиях разной освещенности местопроизрастаний
| Репродуктивные показатели | Высокая освещённость | Затенение | t-критерий Стьюдента |
| Масса 1000 семян, мг, M±m | 8,00±0,01 | 11,53±0,08 | 44,13*** |
| Количество семян в 1 г, шт., M±m | 12376±0,03 | 8680±0,04 | 12,83*** |
| Реальная семенная продуктивность | 2766,10±6,32 | 1354,00±65,62 | 15,33*** |
Примечания: M±m – среднее арифметическое ± ошибка среднего;
различия достоверны при Р≥0,999 (***).
Выявлено, что у семян, собранных на затенённых участках, лабораторная всхожесть в 2 раза ниже чем на незатенённых (21,56 % и 43,50 % соответственно). С уменьшением уровня освещённости местопроизрастания у R. kirilowii достоверно снижается и реальная семенная продуктивность (табл. 1). При этом в затенении масса 1000 семян достоверно больше, а их количество в 1 г меньше. Поэтому для семенного размножения R. kirilowii маточники рекомендуется выращивать на участках с высокой освещённостью.
Поскольку при выращивании из семян молодые растения R. kirilowii декоративны только с третьего года жизни, для зелёного строительства семенное размножение данного вида экономически менее целесообразно, чем размножение методом черенкования.
При укоренении черенков R. kirilowii в условиях защищённого грунта в песке и почвосмеси и сразу в открытом грунте на освещённых и затенённых участках нами отмечена 100 % укореняемость черенков во всех вариантах эксперимента.
При анализе размеров образованной корневой системы выявлено, что способ укоренения не влияет на её длину, а максимального распространения в ширину корневая система достигает в песке в условиях защищённого грунта (табл. 2).
Таблица 2. Размеры корневой системы, образованной при укоренении черенков Rhodiola kirilowii (Regel) Maxim. разными способами
| Размеры корневой системы, см, M±m | В песке | В почвосмеси | В открытом грунте | |
| высокая освещённость | затенение | |||
| Максимальная длина корневой системы | 7,13±0,35 | 7,07±0,65 | 7,01±0,82 | 7,02±0,93 |
| Максимальное распространение корневой системы в ширину | 5,65±0,37 | 4,10±0,40 | 3,96±0,74 | 4,02±0,61 |
Примечания: M±m – среднее арифметическое ± ошибка среднего.
Следовательно, данный вид целесообразнее укоренять сразу на постоянном месте в цветочной композиции, независимо от освещённости будущего местопроизрастания, желательно в почве с добавлением песка.
Выводы и заключение
В течение 7 лет в условиях Донецкого ботанического сада R. kirilowii проходит этапы развития от латентного периода до зрелого генеративного возраста. С третьего по седьмой годы жизни растения декоративны. Плодоношение генеративных особей ежегодное, при этом самосев единичный и нерегулярный. Для семенного размножения R. kirilowii маточники рекомендуется выращивать на участках с высокой освещённостью. При вегетативном размножении данный вид целесообразнее укоренять сразу на постоянном месте в цветочной композиции, независимо от освещённости будущего местопроизрастания, желательно в почве с добавлением песка.
Литература
Бабиченко В. Н., Барабаш М. Б., Логвинов К. Т. и др. Природа Украинской ССР: Климат. Киев: Наук. думка, 1984. 232 с.
Бельгардт А. Л. К вопросу об экологическом анализе и структуре лесных фитоценозов в степи // Вопросы биологической диагностики лесных биогеоценозов Приморья. Днепропетровск, 1980. С. 12—43.
Борисова А. Г. Конспект системы сем. Crassulaceae DC. флоры СССР (добавления и изменения) // Новости систематики высших растений. Л., 1969. Т. 6. С. 112—121.
Вайнагий И. В. О методике изучения семенной продуктивности растений // Ботан. журн. 1974. Т. 59. № 6. С. 826—830.
Воронцова Л. И., Гатцук Л. Е., Егорова В. Н. и др. Ценопопуляции растений (основные понятия и структура). М.: Наука, 1976. 214 с.
Геоботанічне районування Української РСР. К.: Наук. думка, 1977. 302 с.
Зиман С. Н. Жизненные формы и биология степных растений Донбасса. Киев: Наук. думка, 1976. 190 с.
Игнатьева И. П. Методика изучения морфогенеза вегетативных органов травянистых поликарпиков // Докл. ТСХА. 1964. № 98. С. 47—57.
Краснов Е. А., Саратиков А. С., Суров Ю. П. Растения семейства Толстянковых. Томск, 1979. 207 с.
Куперман Ф. М. Исследование закономерностей морфогенеза растений методом выращивания их в условиях разных световых режимов // Свет и морфогенез растений / Под ред. Ф. М. Купермана, Е. И. Ржановской. М.: Изд-во Моск. ун-та, 1978. С. 8—43.
Методические указания по семеноводству интродуцентов / Отв. ред. Н. В. Цицин. М., 1980. 64 с.
Ничипорович А. А., Кефели В. И., Акназаров О. А. Действие световых факторов высокогорий Памира на жизнедеятельность растений. Душанбе: Дониш, 1985. 216 с.
Привалко Л. В. Декоративные толстянковые для использования в ландшафтном фитодизайне: рекомендации / Под общ. ред. А. З. Глухова. Донецк, 2010. 20 с.
Работнов Т. А. Методы изучения семенного размножения травянистых растений в сообществах // Полевая геоботаника. М., Л.: Наука, 1960. Т. 2. С. 20—40.
Работнов Т. А. Определение возрастного состава популяций видов в сообществе // Полевая геоботаника. М., Л.: Наука, 1964. Т. 3. С. 132—208.
Работнов Т. А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Тр. БИН АН СССР. 1965. Сер. 3. № 6. С. 7—204.
Серебряков И. Г. Экологическая морфология растений. Жизненные формы покрытосеменных и хвойных. М.: Высш. шк., 1962. 378 с.
Смирнова О. В., Заугольнова Л. Б., Торонова Н. А. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф // Ценопопуляция растений. М.: Наука, 1976. С. 14—43.
Тарасов В. В. Флора Дніпропетровської та Запорізької областей. Судинні рослини. Біолого-екологічна характеристика видів. Д.: Вид-во ДНУ, 2005. 276 с.
Уранов А. А. Жизненные состояния вида в растительном сообществе // Бюлл. МОИП. Сер. Биол. 1960. Вып. 3. С. 77—92.
Флора Западной Сибири. Томск, 1964. Т. 12. 327 с.
Флора Казахстана: в 9 т. Алма-Ата, 1961. Т. 4. 344 с.
Raunkiaer C. Planterigets Livsformer of deres Betydning for Geografien. Kobenhavn: Nordiskforland, 1907. 132 p.
The Plant List, 2013. Version 2; URL: http://www.theplantlist.org/ (дата обращения 20.02.2017).
