

Спорово-пыльцевой анализ используется для палеоклиматических и палеоэкологических реконструкций, необходимых для понимания процессов, происходящих в климатической системе планеты в настоящем и их развития в будущем. Изучение спорово-пыльцевых спектров с привлечением данных об окружающей места сбора растительности показывает особенности формирования воздушных и поверхностных спектров, а также уточняет как состав спектров отражает продуцирующую их растительность.
Представления о закономерностях распространения пыльцы и спор в пространстве начали формироваться ещё в первые этапы развития палинологических исследований. Сведения о дальности распространения пыльцы наиболее часто встречающихся таксонов представлены у А. Н. Сладкова (1967). Один из самых подробных детальных обзоров методов изучения репрезентативности пыльцевых спектров был сделан Т. Giesecke et al. (2010). М. Б. Носовой (2020) проведена ревизия методических работ по изучению воздушных пыльцевых спектров, а также методов обработки и интерпретации данных, полученных с помощью спорово-пыльцевого анализа. Одной из первых С. Г. Губанкова (1981) начала аэропалинологические исследования на территории современной России. В воздухе Ленинграда мониторинг аллергеногенной пыльцы проводили Л. Г. Никольская и Г. Б. Федосеев (1987). Пыльцевые аэрозоли воздушных масс юга Западной Сибири исследовал В. В. Головко (2001). В Европейской части России также проводились подобные исследования, например, О. В. Морозовой и Р. Я. Мешковой (2006) были получены данные о пыльцевом дожде для Смоленской области, Ю. М. Посевина (2011) изучала динамику поллинозов в зависимости от пыления разных аллергеногенных растений в Рязани. Исследования отображения различных биомов средней части Восточно-Европейской равнины (от южной тайги до лесостепи) в пыльцевых спектрах были предприняты на территориях Полистовского государственного природного заповедника в Псковской области (Nosova et al., 2019), Центрально-Лесного государственного природного биосферного заповедника в Тверской области (Nosova et al., 2012), Звенигородской биостанции им. С. Н. Скадовского Биологического факультета МГУ им. М. В. Ломоносова в Московской области, Тульских засек и музея-заповедника «Куликово поле» в Тульской области (Новенко и др., 2011; Носова, 2009; Nosova et al., 2015, 2020; Severova, Volkova, 2017). В работе Lisitsyna et al. (2017) представлены результаты исследования воздушных спектров в Республике Коми (Северо-восток Европейской части России). Воздушные спектры и репрезентативность древесной растительности изучали в пыльцевых спектрах на севере Финляндии – в Лапландии (Huusko, Hicks, 2009; Hicks, Hyvärinen, 2014).
Первые исследования воздушных спектров Петрозаводска (Республика Карелия) были предприняты Н. А. Елькиной и Е. Ф. Марковской (2007). В целях создания календаря пыления для территории Петрозаводска за период с 2009 по 2016 года Н. А. Елькиной (2018) были изучены воздушные спектры селитебной зоны Петрозаводска. При этом пыльцевые спектры территорий, окружающих город Петрозаводск, не изучались. С этой целью нами была выбрана территория Ботанического сада ПетрГУ, для которой точно установлена интродуцированная и автохтонная, занимающая заповедную территорию, растительность.
Петрозаводск - столица Республики Карелия на Северо-Западе Российской Федерации. Город расположен на северо-западе Онежского озера, на берегу Петрозаводской Губы. Ботанический сад Петрозаводского Государственного Университета находится, примерно в восьми километрах к северу-северу-востоку от центра Петрозаводска, на северном побережье Петрозаводской Губы Онежского озера (Рис. 1). Западная граница Ботанического сада отделяет его от селитебной территории района Соломенное города Петрозаводск, занимающего оба берега протоки, соединяющей озеро Логмозеро и Петрозаводскую Губу Онежского озера. Площадь Ботанического сада составляет 367 га. Сад заложен в 1951 году (Сайт Ботанического …, 2023).
Особое внимание обращалось на пыльцу семейства Ореховых (Juglandaceae), выбранное в качестве модельного объекта из-за хорошей распознаваемости пыльцы, своей экзотичности для таёжной зоны и, одновременно, присутствия Ореха манчжурского (Juglans mandschurica Maxim.) в экспозиции Ботанического сада ПетрГУ. В Ботаническом саду произрастают девять деревьев представителей Ореховых, семь из которых цветут. Семейство Ореховых представляет интерес не только как экономически значимое семейство, но и как семейство, к которому принадлежат красивые декоративные растения, что могло бы иметь потенциал для зелёного хозяйства (Kozlowski et al., 2018).
До сих пор точно неизвестно, как разные типы пыльцы и какое количество зёрен отображают разные типы растительного покрова в пыльцевых спектрах; как влияет на этот процесс продуктивность пыльцы; на какое расстояние могут распространяться пыльцевые зёрна разных типов от продуцирующей их растительности и как разные типы почв влияют на сохранность пыльцевых зёрен.
Цель данной работы – выявить, как растения разных таксонов представлены в пыльцевых спектрах Ботанического сада ПетрГУ и города Петрозаводска для пыльцевого прогноза и палеогеографических и климатических реконструкций.
Отдельной целью данного исследования являлось определение в пыльцевых спектрах интродуцентов, в частности, представителя семейства Ореховых (Juglandaceae) – Ореха манчжурского (J. mandschurica), произрастающего на территории Ботанического сада ПетрГУ.
Для ответа на перечисленные вопросы были поставлены следующие задачи: 1) установить пыльцевые ловушки на вегетационный период и изучить пыльцевые спектры; 2) исследовать субрецентные пыльцевые спектры в местах установки ловушек на территории Ботанического сада ПетрГУ и на отдалении в городе Петрозаводске; 3) проанализировать полученные данные и оценить репрезентацию интродуцентов и нативной растительности исследуемых участков в пыльцевых спектрах.
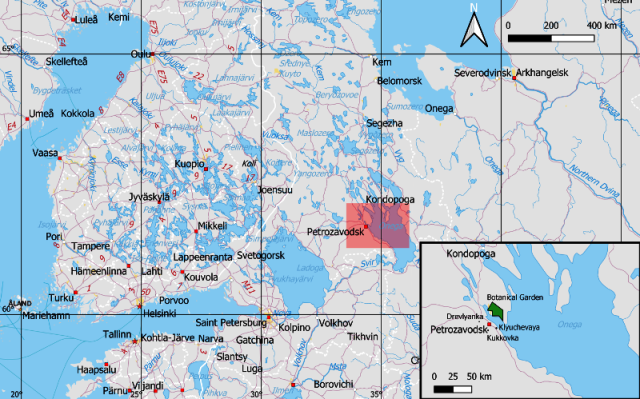
Рис. 1. Обзорная карта исследуемой территории (создана в QGIS, с применением Natural Earth Data).
Fig. 1. Study area map (made in QGIS with Natural Earth Data).
Ботанический сад ПетрГУ занимает северное побережье Петрозаводской Губы Онежского озера, на меньшей части производится интродукция и акклиматизация растений, а на большей части - участок заповедной территории с ненарушенным естественным покровом средней тайги. Тестовые площадки были расположены в разных частях Ботанического сада (Рис. 2), как на окультуренных участках, так и на территории охраняемых природных ландшафтов. Для исследования пыльцевого дождя нами были установлены восемь пыльцевых ловушек гравитационного типа на два вегетационных сезона 2019 и 2020 годов. Ловушки, пятилитровые пластиковые бутыли с горлышком пять сантиметров в диаметре, фиксировались, согласно несколько модифицированной нами методике, описанной в работе Х. Таубера (Tauber, 1974), на высоте 1.5 м над земной поверхностью, на стволах следующих интродуцентов: Hippophaё rhamnoides L. (61°78′87.71″N 34°38′23.56″E), Pseudotsuga menziesii Franco (61°84′29.79″N 34°38′99.81″E), Juglans mandschurica (61°84′32.33″N 34°38′89.49″E), Larix sibirica Ledeb. (61°84′36.95″N 34°38′45.69″E), Fraxinus excelsior L. (61°84′47.77″N 34°38′86.87″E), Ulmus laevis Pall. (61°84′25.49″N 34°38′72.38″E), Quercus rubra L. (61°84′22.30″N 34°38′90.88″E), а также деревьев, произрастающих в естественном ненарушенном таёжном биоме (61°84′24.59″N 34°37′98.24″E). В местах инсталяции пыльцевых ловушек были отобраны образцы дернин для исследования субрецентных спорово-пыльцевых спектров (ТС1 - Европейский сектор, Acer; ТС2 - Граница экспозиции, Pinetum-uliginosum; ТС3, ТС3а - Европейский сектор, Ulmus; ТС4 - Между прудами, Tilia; ТС5, ТС5а - Американский сектор, Pseudotsuga; ТС6, ТС6а - У администрации; ТС7, ТС7а - Азиатский сектор, Juglans; ТС8 – Quercus; ТС9 - Плодовый сад, Hippophaë; ТС10 – Larix; ТС11 – Fraxinus). Также были отобраны образцы дернин из центра Петрозаводска (ТС12 – пересечение пр. Ленина и ул. Фридриха Энгельса; ТС13 – у ж/д вокзала) и на расстоянии 10 км от Ботанического сада ПетрГУ (ТС14 – парк Кукковка; ТС15 – ул. Ключевая; ТС16 – микрорайон Древлянка) Нами были выбраны следующие хорошо определяемые палинологические маркеры древесных интродуцентов: P.menziesii, J. mandshurica, A. hippocastanum, U. laevis, Q. rubra, Elaeagnus commutata Bernh. ex Rydb.. На пыльцу пограничных, чаще интродуцированных видов, таких как Acer platanoides L. и Tilia cordata Mill., вблизи Петрозаводска находится северная граница ареалов их произрастания), тоже обращалось пристальное внимание.
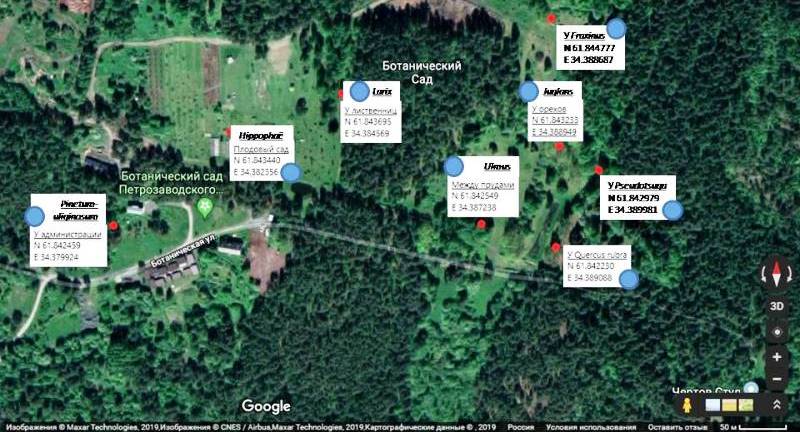
Рис. 2. Карта мест установки пыльцевых ловушек (маркировка синим цветом) и взятия почвенных проб (красным) на территории Ботаничсекого сада ПетрГУ (создана в Google Maps).
Fig. 2. Map of sampling sites in the Petrozavodsk Botanical Garden. In red colour – soil sampling sites and in blue colour – pollen traps installation sites are indicated (made in Google Maps).
Для исследования субрецентных спорово-пыльцевых спектров были отобраны 15 почвенных проб (дернина) на территории Ботанического сада, из них восемь непосредственно в местах установки пыльцевых ловушек. Также были отобраны пять почвенных проб из мест, расположенных в радиусе 10 км от Ботанического сада, на территории города Петрозаводск (Рис. 3): в сквере на пересечении проспекта Ленина и улицы Фридриха Энгельса (у гостиницы «Северная») (61°78′87.71″N 34°36′01.03″E), в сквере у железнодорожного вокзала (61°78′46.52″N 34°34′60.97″E), в районе «Ключевая» (61°76′ 99.98″N 34°43′26.85″E), в парке «Кукковка» (61°76′24.22″N 34°38′18.05″E) и на границе города в районе «Древлянка» (61°75′67.97″N 34°29′12.73″E). Всего за два года было исследовано 36 проб воздушных и почвенных спектров.
Содержимое использованных в течении сезона ловушек переливали в пробирки, ловушки промывали дистиллированной водой, центрифугировали жидкости в пробирках, сливали воду, оставляя на дне пробирок твердый осадок. Далее твердый осадок из пыльцевых ловушек обрабатывался по ацетолизному методу Г. Эрдтмана (Erdtman, 1943). Почвенные пробы подвергались обработке по общепринятой методике В. П. Гричука (Гричук, Заклинская, 1950). Для микроскопирования были приготовлены постоянные препараты на глицерин-желатиновой основе. Препараты изучали с помощью светового микроскопа Микмед-6 при увеличении в 400 раз. Микрофотографирование осуществлялось при увеличении в 1000 раз с применением нефлюоресцирующего иммерсионного масла МиниМед. Для идентификации пыльцевых зёрен использовались Атласы пыльцы и спор (Куприянова, Алёшина, 1972; 1978), материалы коллекции споротеки лаборатории палинологии БИН РАН, коллекция референсной пыльцы, созданной авторами.
Пыльцевые диаграммы были выполнены с помощью программы C2 по методике, описанной S. Juggins (2007).
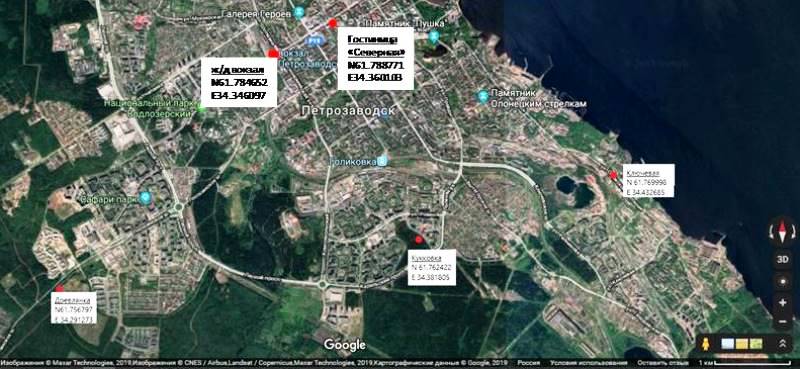
Fig. 3. Map of soil sampling sites in Petrozavodsk (made in Google Maps).
Во всех полученных пыльцевых спектрах хорошо представлена региональная растительность, в данном случае, средней тайги. Повсеместно обнаружены пыльцевые зёрна таких древесных растений как Pinus, Picea, Alnus, Betula. Реже встречаются пыльцевые зёрна Salix и различных травянистых растений из семейств Poaceae, Asteraceae, Apiaceae, Brassicaceae, Cyperaceae и других.
Пыльца представителей интродуцируемой растительности лучше всего представлена в ловушках, непосредственно расположенных на самих экзотах. Пыльцевые зёрна некоторые неместных растений были также найдены в спектрах ловушек, расположенных на заповедной территории. В результате исследования в пыльцевых спектрах выявлена пыльца следующих древесных интродуцентов: из голосеменных – Pseudotsuga menziesii, из покрытосеменных – представителей из родов Ulmus, Quercus, Juglans, Corylus, Syringa, Fraxinus, Berberis, Morus а также представителей произрастающих на северной границе своих ареалов распространения Acer platanoides и Tilia cordata. Пыльца изучаемых интродуцентных видов Aesculus hippocastanum и Elaeagnus commutata в палиномах не обнаружена.
Суммарно в спектрах определена пыльца 46 таксонов из 38 семейств, в том числе шести голосеменных и 40 покрытосеменных, и три типа спор (Табл. 1).
Анализ количественного и качественного состава спорово-пыльцевых спектров представлен на пыльцевых диаграммах (Рис. 4-5).
Таблица 1. Таксоны и встречаемость их пыльцы и спор в почвенных и воздушных пробах.
Table 1. Pollen and spores’ taxa and their occurrence in soil and aerial samples.
| № | Таксон Taxa | Число почвенных проб, содержащих пыльцу таксона Occurrence in soil samples | Число воздушных проб, содержащих пыльцу таксона Occurrence in aerial samples |
| Древесные растения Arboreal | |||
| Голосеменные Conifers | |||
| 1 | Pinus | 20 | 16 |
| 2 | Picea | 18 | 16 |
| 3 | Abies | 0 | 8 |
| 4 | Larix | 12 | 10 |
| 5 | Pseudotsuga | 6 | 2 |
| 6 | Taxaceae/Cupressaceae | 0 | 2 |
| Цветковые Angiosperms | |||
| Лиственные Deciduous | |||
| 1 | Alnus | 19 | 16 |
| 2 | Betula | 20 | 16 |
| 3 | Corylus | 6 | 7 |
| 4 | Acer | 2 | 1 |
| 5 | Cornus | 1 | 0 |
| 6 | Tilia | 9 | 4 |
| 7 | Ulmus | 9 | 6 |
| 8 | Quercus | 3 | 12 |
| 9 | Morus | 1 | 0 |
| 10 | Juglans | 7 | 12 |
| 11 | Oleaceae (Syringa) | 4 | 1 |
| 12 | Fraxinus | 0 | 3 |
| 13 | Rosaceae | 2 | 10 |
| 14 | Salix | 1 | 4 |
| 15 | Berberis | 0 | 1 |
| Кустарники Shrubs | |||
| 1 | Ribes | 1 | 0 |
| 2 | Ericaceae (Rhododendron) | 3 | 2 |
| 3 | Adoxaceae (Virbunum) | 1 | 1 |
| 4 | Hippophaë | 0 | 1 |
| Травянистые растения Herbaceous | |||
| 1 | Poaceae | 20 | 15 |
| 2 | Однобороздные однодольные Moncolpate Monocots | 10 | 1 |
| 3 | Asteraceae | 15 | 4 |
| 4 | Cichorium | 9 | 0 |
| 5 | Apiaceae | 11 | 5 |
| 6 | Lamiaceae | 4 | 2 |
| 7 | Ranunculaceae | 2 | 2 |
| 8 | Urtica | 1 | 0 |
| 9 | Brassicaceae | 4 | 9 |
| 10 | Caryophyllaceae | 2 | 2 |
| 11 | Chenopodiaceae | 7 | 1 |
| 12 | Cyperaceae | 6 | 5 |
| 13 | Malvaceae | 1 | 1 |
| 14 | Dipsacus | 1 | 0 |
| 15 | Polygonaceae | 1 | 0 |
| 16 | Drosera | 1 | 0 |
| 17 | Fabaceae | 0 | 1 |
| 18 | Geranium | 0 | 1 |
| 19 | Campanulaceae | 0 | 2 |
| 20 | Onagraceae | 0 | 2 |
| 21 | Fumaria | 0 | 1 |
| Споровые растения Sporogenous | |||
| 1 | Polypodiaceae | 19 | 6 |
| 2 | Трёхлучевые Triletes | 18 | 1 |
| 3 | Lycopodium | 1 | 0 |
Воздушные спектры
Практически во всех образцах, отобранных в 2019 году (Рис. 4), кроме ловушки в Ботаническом саду на краю леса, обнаружены пыльцевые зёрна древесных интродуцентов, содержанием менее одного-двух процентов. В отдельных ловушках выявляется до 33 процентов пыльцы видов, на которых расположены ловушки. Почти в половине образцов из ловушек (11 из 23) из Ботанического сада представлены зёрна двух различных представителей древесных экзотов, в других же встречается пыльца одного (в трёх образцах), трёх (в двух образцах), четырёх (в трёх образцах), пяти (в двух образцах) и шести (в одном образце), представителей деревьев и кустарников интродуцентов (Рис. 4-5). Таким образом, разнообразие пыльцевых зёрен в ловушках невелико, ограничено произрастающими поблизости деревьями.
В воздушных спектрах, собранных за 2020 год выявлены следующие особенности, показанные на диаграмме (Рис. 4). Увеличение доли представителей Poaceae почти во всех пыльцевых спектрах не более одного процента относительно 2019 года. Рост доли пыльцы Alnus в пыльцевых спектрах в среднем на пять процентов, за исключением ловушек на Juglans и Pseudotsuga, где отмечено уменьшение вклада пыльцы ольхи (Alnus) в формирование пыльцевых спектров на 25 и пять процентов соответственно. Отмечается уменьшение роли Betula в пыльцевых спектрах в среднем на пять процентов, за исключением ловушек на Juglans и у администрации, где, наоборот, отмечен рост процентного содержания берёз (Betula) на пять процентов. Заметно увеличение количества пыльцы Pinus в большинстве спектрах ловушек от пяти до 25 процентов, хотя в трёх ловушках (на вязе (Ulmus), у администрации и на дубе (Quercus)) отмечается снижение количества пыльцы этого таксона. Важно отметить, что пыльца Picea в материалах 2020 года представлена намного чаще и на два-пять процентов больше, чем в образцах 2019 года. Характерно резкое увеличение доли пыльцы Ulmus и Quercus в ловушках, находящихся на «своих» деревьях, на 20 и на 30 процентов соответственно. При этом пыльца Juglans и Pseudotsuga почти исчезлa из пыльцевых спектров в 2020 году – доли обоих таксонов стали составлять меньше процента. Пыльца таких древесных таксонов как Salix и Corylus тоже имеет лучшую репрезентацию в спектрах 2020 года. Из пыльцы представителей травянистой растительности, обнаруженной более широко в пыльцевых спектрах 2020 года по сравнению с 2019 годом, можно отметить пыльцу таких таксонов как Cyperaceae, Brassicaceae, Apiaceae, Ranunculaceae. Все они составляют долю меньше одного процента в пыльцевых спектрах ловушек.
Воздушные спектры, изученные за 2019 и 2020 года, характеризуются наличием пыльцы древесной растительности, а в особенности, пыльцой тех растений, на которые были установлены пыльцевые ловушки. Во всех спектрах большую долю составляют пыльцевые зёрна, репрезентирующие региональный компонент спектров: Pinus и Betula, и в меньшем количестве Picea и Alnus. Пыльца трав составила очень малу долю изученных спектров (Рис. 4).
Почвенные спектры
Во всех почвенных пробах доминирует пыльца растений древесных таксонов, её содержание в образцах из Ботанического сада (Рис. 5) составляет 80 – 98 процентов, только в пробе ТС11 древесных таксонов меньше, около 60 процентов, но в ней больше всего обнаружено спор (около 20 процентов). В городских пробах (Рис. 5) древесная растительность представлена в чуть меньшем количестве, от 68 до 90 процентов. В палиномах Ботанического сада значительно преобладает пыльца хвойных от 48 до 93 процента, причём главным образом за счёт пыльцевых зёрен сосны (Pinus), которые встречаются во всех исследованных спектрах в количестве 35 – 91 процент от общего числа пыльцы и спор). Во всех субрецентных пробах из Ботанического сада и в трёх из пяти городских проб доминирует пыльца Pinus. В городе процент пыльцы хвойных несколько уменьшается от 42 до 81 процента, как и сосновой пыльцы (28 – 65 процентов). Также во всех спектрах, кроме образца ТС6а, обнаружены пыльцевые зёрна ели (Picea), однако в существенно меньшем числе. В Ботаническом саду их количество составляет 1.2 – 15 процентов, а в городе увеличивается до 8 – 38 процентов. Максимальное количество пыльцы Picea в пробе (ТС14) из ельника.
Пыльца пихты (Abies) зафиксирована в единичном экземпляре только в одной пробе в Ботаническом саду в ловушке, а лиственница (Larix) обнаруживалась регулярно, но не превышая один-два процента. Taxaceae/Cupressaceae характерны только для восьми из 28 проб, содержание пыльцы растений этих таксонов варьирует от 0.2 до 6.4 процентов. Пыльца единственного хвойного древесного интродуцента, псевдотсуги (Pseudotsuga), найдена вблизи дерева в незначительном количестве (0.8 процента).
Из пыльцы древесных цветковых в почвенных пробах обнаружены пыльцевые зёрна 15 родов из 11 семейств, большинство из которых являются интродуцентами для Петрозаводска. Пыльца берёз (Betula) в субрецентных образцах составляет от 0,7 до 21 процента, а ольхи (Alnus) от 0.3 до 9.5 процентов палинома. В одной городской пробе (ТС16) доминирует пыльца Betula (36.9 процентов). За исключением представителей Betulaceae, всегда характеризующих субрецентные спектры Петрозаводска, остальные древесные скудно представлены от одного до двух процентов спорово-пыльцевых спектров.
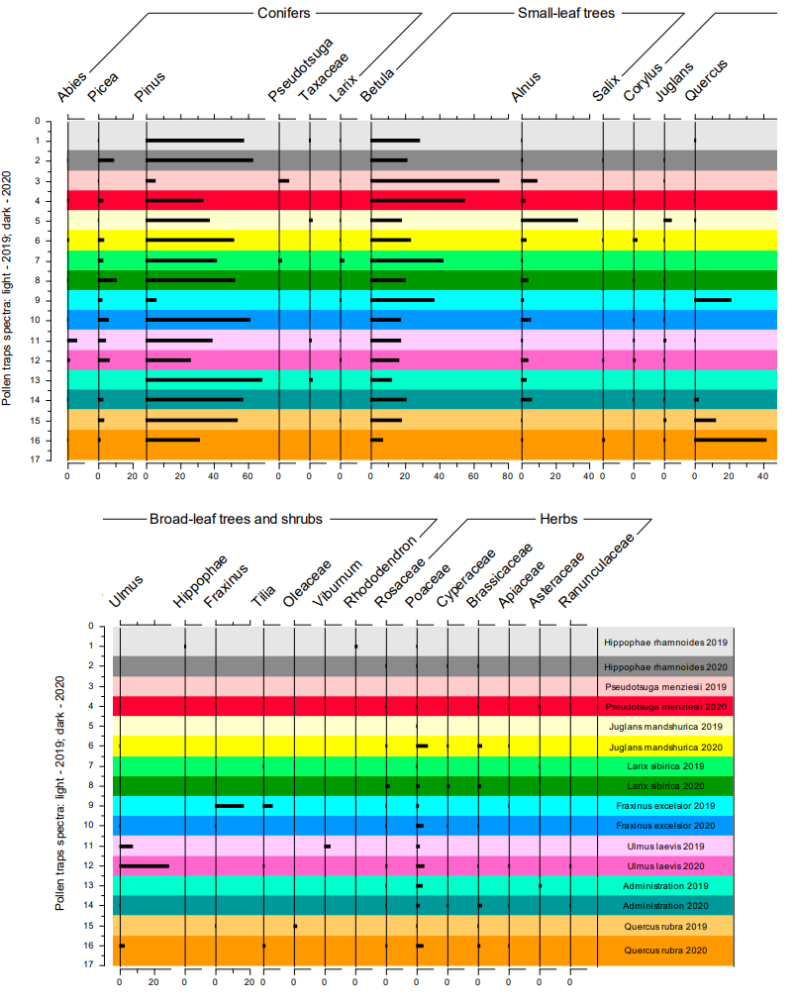
Рис. 4. Пыльцевая диаграмма воздушных спектров вегетационных сезонов 2019 и 2020 годов Ботанического сада ПетрГУ (создана с использованием программного обеспечения C2).
Fig. 4. Pollen diagram of aerial spectra of 2019 and 2020 years seasons from the Petrozavodsk State University Botanic garden (made with C2 software).
Травянистые растения в спектрах представлены незначительно (0.3 – 7.5 процентов). Постоянным компонентом палинома являются ветроопыляемые Злаки (Poaceae). В зоне средней тайги количество их пыльцы в субрецентных пробах варьирует от 0.1 до 3.2 процента. На втором месте по частоте встречаемости представителей семейств идут Сложноцветные (Asteraceae) (включая тип Цикорий (Cichorium)), их пыльца обнаружена в 15 из 20 исследованных субрецентных проб. Процентное содержание пыльцы Астровых (Asteraceae), как и всех трав, в поверхностных пробах лесной зоны незначительно, обычно менее одного процента, редко достигая четырёх – шести процентов. Некоторое увеличение числа пыльцевых зёрен Сложноцветных (Asteraceae) или наличие зёрен других конкретных таксонов трав вероятно связано с тем, что проба взята непосредственно вблизи произрастания и цветения этих растений. В 10 из 20 субрецентных проб выявлены однобороздные пыльцевые зёрна с сетчатой или перфорированной поверхностью. В условиях Петрозаводска эти зёрна следует относить к травянистым однодольным. Встречаются единичные пыльцевые зёрна представителей еще 12 семейств: Зонтичных (Apiaceae), Губоцветных (Lamiaceae), Лютиковых (Ranunculaceae), Крапивных (Urticaceae), Крестоцветных (Brassicaceae), Гвоздичных (Caryophillaceae), Маревых (Chenopodiaceae), Осоковых (Cyperaceae), Мальвовых (Malvaceae), Гречишных (Polygonaceae), Ворсянковых (Dipsacaceae), Росянковых (Droseraceae). Чаще других в субрецентных пробах обнаруживаются единичные зёрна Зонтичных (Apiaceae) и Губоцветных (Lamiaceae).
Во всех почвенных пробах выявлены споры высших растений в количестве от 0.3 до 19.1 процентов.
В городе помимо двух проб без пыльцы интродуцентов ещё в трёх пробах показано наличие зёрен широко культивируемых в городских условиях сирени (Syringa), вяза (Ulmus), лещины (Corylus), ореха (Juglans) и липы (Tilia) по два-четыре таксона на пробу, но общее содержание их не превышает два процента.
В Петрозаводске выявлена пыльца растений шести таксонов голосеменных из семейств Pinaceae (пять родов – Pinus, Picea, Abies, Larix, Pseudotsuga) и плохо отделяемых друг от друга Taxaceae/Cupressaceae. Среди древесных покрытосеменных встречаются пыльцевые зёрна представителей 12 семейств Betulaceae (родов Alnus, Betula, Corylus), Aceraceae (Acer), Cornaceae (Cornus), Tiliaceae (Tilia), Ulmaceae (Ulmus), Fagaceae (Quercus), Juglandaceae (Juglans), Moraceae (Morus), Berberidaceae (Berberis), Oleaceae (Syringa, Fraxinus), Salicaceae (Salix) и Rosaceae. Практически все выявленные в пыльцевых спектрах цветковые древесные растения, кроме Alnus, Betula и некоторых розоцветных (Rosaceae), являются интродуцентами.
Сравнение рецентных и субрецентных спорово-пыльцевых спектров
(из ловушек и почв)
Как и в субрецентных пробах, во всех пыльцевых ловушках господствует пыльца древесных растений (до 98 процентов). При анализе встречаемости пыльцы отдельных представителей обнаруживаются различия содержания пыльцевых зерен (Рис. 4-5). Так, выявлено, что не во всех спектрах доминируют голосеменные. Так, в образцах двух из 16 ловушек пыльца хвойных составляет 10-12 процентов, в восьми ловушках хвойные представлены в количестве 40 – 60 процентов, а в ловушке, установленной у администрации, прикреплённой к сосне (Pinus), пыльца хвойных составляет 70.7 процентов, из них пыльца сосны - 68.5 процентов. Пыльцевые зёрна сосны присутствуют в спектрах всех ловушек, но в меньших количествах, чем в субрецентных пробах. В двух ловушках с наименьшим содержанием пыльцы хвойных (10.7 – 12.8 процентов) Pinus составляет примерно половину спектра хвойных, 5.6 – 6.2 процентов. В пяти ловушках количество зёрен Pinus колеблется от 37.7 до 57.7 процентов всего палинома, что является значительной долей пыльцевого спектра голосеменных конкретной ловушки. Во всех ловушках обнаружена пыльца ели (Picea) в среднем меньше, чем в почвенных пробах, от 0.2 до 9.7 процентов. Пыльца псевдотсуги (Pseudotsuga) в количестве 6.11 процентов выявлена в ловушке, прикреплённой к этому дереву, ещё в одной ловушке её содержание составляет около двух процентов. Также в ловушках постоянно присутствуют пыльцевые зёрна лиственниц (Larix) и/или представителей Taxaceae/Cupressaceae в размахе от 0.3 до 2.7 процентов (Рис. 6: 3). Пыльца Taxaceae/Cupressaceae была обнаружена в четверти образцов воздушных спектров (четыре из 16 проб) и в одной четверти образцов почвенных спектров (пять из 20 проб).

Рис. 5. Пыльцевая диаграмма субрецентных (почвенных) спектров Ботанического сада ПетрГУ (ТС1-ТС11) и города Петрозаводска (ТС12-ТС16) (создана с использованием программного обеспечения C2).
Условные обозначения: ТС1 - Европейский сектор, Acer; ТС2 - Граница экспозиции, Pinetum-uliginosum; ТС3, ТС3а - Европейский сектор, Ulmus; ТС4 - Между прудами, Tilia; ТС5, ТС5а - Американский сектор, Pseudotsuga; ТС6, ТС6а - У администрации; ТС7, ТС7а - Азиатский сектор, Juglans; ТС8 – Quercus; ТС9 - Плодовый сад, Hippophaë; ТС10 – Larix; ТС11 – Fraxinus; ТС12 – пересечение пр. Ленина и ул. Фридриха Энгельса; ТС13 – у ж/д вокзала; ТС14 – парк Кукковка; ТС15 – ул. Ключевая; ТС16 – микрорайон Древлянка.
Fig. 5. Pollen diagram of subfossil (soil) spectra from the Petrozavodsk State University Botanic garden (TC1-TC111) and Petrozavodsk (TC12-TC16) (made with C2 software).
Legend: TC1 – European sector, Acer; TC2 – Border of the exposition, Pinetum-uliginosum; TC3, TC3a – European sector, Ulmus; TC4 – Between the ponds, Tilia; TC5, TC5a – American sector, Pseudotsuga; TC6, TC6a – Administration; TC7, TC7a – Asian sector, Juglans; TC8 – Quercus; TC9 - Horticultural garden, Hippophaë; TC10 – Larix; TC11 – Fraxinus; TC12 – Lenin Avenue – Friedrich Engels Street cross; TC13 – Railway station; TC14 – Kukkovka Park; TC15 – Klyuchevaya Street; TC16 – Drevlyanka District.
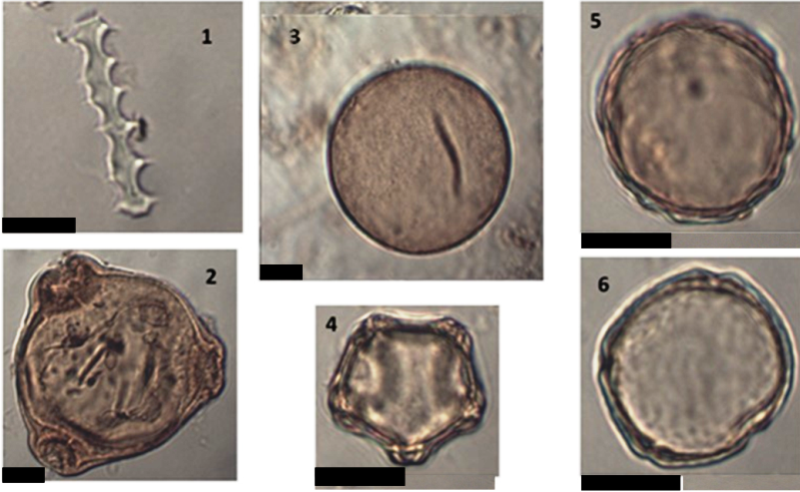
Рис. 6. Микрофотографии некоторых пыльцевых зёрен и непыльцевых палиноморф, найденных в спектрах пыльцевых ловушек, установленных в Ботаническом саду ПетрГУ в 2020 году. Все микрофотографии сделаны при увеличении в 1000 раз (размерная шкала на фото 1µm).
Условные обозначения: 1 – Остаток (фитолит) Poaceae, пыльцевая ловушка на Hippophaё; 2 - Пыльцевое зерно Chamaenerion, пыльцевая ловушка на Betula рядом с Pseudotsuga; 3 - Пыльцевое зерно Larix, пыльцевая ловушка на Larix; 4 - Пыльцевое зерно Alnus, пыльцевая ловушка на Larix; 5 - Пыльцевое зерно Juglans, пыльцевая ловушка на Quercus; 6 - Пыльцевое зерно Ulmus, пыльцевая ловушка на Ulmus
Fig. 6. Microphotographs of some pollen taxa and non-pollen palynomorphs found in pollen traps spectra from the Petrozavodsk State University Botanic Garden in 2020. All microphotographs are made with magnification 1000 times (Scale bar 1µm)
Legend: 1 - Poaceae remain (phytolith), pollen trap on the Hippophaё; 2 - Chamaenerion pollen grain, pollen trap on the Betula near Pseudotsuga; 3 - Larix pollen grain, pollen trap on the Larix; 4 - Alnus pollen grain, pollen trap on the Larix; 5 - Juglans pollen grain, pollen trap on the Quercus; 6 - Ulmus pollen grain, pollen trap on the Ulmus
В семи из 16 ловушек доминирует пыльца древесных цветковых растений, её содержание составляет 85.3 (ТС5а), 57.2 (ТС7а) и 84.88 (ТС11) процентов. Во всех ловушках выявлен больший процент пыльцы мелколиственных нативных Берёзовых (Betulaceae), главным образом берёзы (Betula) - от 12.6 до 75 процентов в 16 ловушках, тогда как в 16 субрецентных пробах от 0.7 до 11.2 процентов. Увеличенное содержание Betula отмечено только в двух пробах (до 14 и 21 процента) из Ботанического сада и в двух (до 21 и 37 процентов) из города. Пыльца берёз господствует в трёх ловушках ТС5а (75 процентов), ТС10 (42.8 процента), и ТС11 (37.2 процентов). Во всех пыльцевых ловушках, так же как и в почвенных пробах, наличие пыльцы древесных интродуцентов обычно довольно низкое (обычно менее одного процента). На содержание пыльцы в ловушках больше влияет её расположение: либо на дереве (интродуценте), либо в непосредственной близости от него.
За 2019 год в ловушке на вязе (Ulmus) выявлено 7.69 процентов пыльцы вяза Ulmus (Рис. 6: 6), в ловушке на орехе (Juglans) – 4.52 процента пыльцы ореха Juglans (Рис. 6: 5), в ловушке на дубе (Quercus) – 12.6 процентов пыльцы дуба Quercus , а в ловушке на ясене (Fraxinus) – 17.05 процентов пыльцы ясеня. Но в этом же спектре ловушки на ясене были ещё представлены близко расположенные к точке сбора пыльцевой материал липы (Tilia) (6.2 процента) и дуба Quercus (21.71 процент). Привнесение пыльцы отмечено на расстоянии не более чем на 10-15 метров от одиночного дерева. Однако и в этих спектрах пыльца древесных интродуцентов не господствует и доминантами остаются сосны Picea и берёзы Betula.
За 2020 год в ловушке на вязе (Ulmus) выявлено 27.67 процентов пыльцы вяза (Рис. 6: 6), в ловушке на орехе (Juglans) – 0.02 процента пыльцы ореха (Рис. 6: 5), в ловушке на дубе (Quercus) – 41.6 процент пыльцы дуба, а в ловушке на ясене (Fraxinus) – 0.01 процента пыльцы ясеня.
Пыльца липы Tilia встречается примерно в половине образцов - в девяти из 20 почвенных проб и в четырёх ловушках – в минимальных количествах, не превышая один процент. Только в ловушке на ясене (Fraxinus) содержание пыльцы Tilia достигает 6,2 процентов в 2019 году. Пыльца Ulmus обнаруживается в спектрах девяти из 20 почвенных проб единичными зёрнами (0.1 – 0.8 процента) и только в одной ловушке – ловушке на этом дереве – выявлено 7.7 процентов пыльцы вяза (Ulmus) в 2019 году и почти 27 процентов в 2020 году. Содержание пыльцы ореха манчжурского (J. mandschurica) отмечено в восьми почвенных пробах и 12 ловушках в пределах 1.7 процента, только в спектре 2019 года было найдено 4.5 процента пыльцы растений данного таксона. Morus, Berberis и Cornus представлены единичными зёрнами (0.1 – 0.3 процента) в одном образце каждый. Пыльца американского дуба красного (Quercus rubra) в спектрах мало отличима от европейского дуба черешчатого (Q. robur L.), обычно произрастающего чуть южнее Санкт-Петербурга, но интродуцируемого в Петрозаводске. Пыльца дубов (Quercus) обнаружена в трёх почвенных пробах и 12 ловушках на территории Ботанического сада. Её содержание в спектрах, в основном, составляет 1.5 процента. Только в ловушках на дубе и вблизи дубов содержание пыльцевых зёрен дуба (Quercus) составляет 12.6 процентов в 2019 и 41.6 процент в 2020 году и 21.71 процент в спектре ловушки на ясене (Fraxinus) за 2019 год. Пыльца сирени (Syringa) чаще встречается в почвенных пробах в городе – в трёх из пяти образцов против двух из 11 образцов в саду, но её количество не превышает двух процентов. В воздушных спектрах она представлена только в спектре 2020 года ловушки на вязе (Ulmus) в малом количестве (0.1 процента). Пыльца ясеня (Fraxinus) обнаружена только в трёх ловушках в саду – в ловушке непосредственно на ясене (Fraxinus) – в оба года (17 процентов в 2019 году и 0.1 процента в 2020 году), и в ловушке на дубе (Quercus) в 2019 году (0.1 процента).
В двух ловушках, также как и в трёх близкорасположенных почвенных пробах, обнаружены зёрна кустарникового интродуцента из семейства Вересковых (Ericaceae) типа Рододендрон (Rhododendron), содержание пыльцы не превышает 0.2 процента. Пыльца облепихи (Hippophaё) выявлена только в ловушке на самом кустарнике в количестве менее одного процента.
Травы во всех ловушках, как и в субрецентных пробах представлены злаками в количестве не более 3.5 процентов от спорово-пыльцевых спектров. Содержание всех травянистых также не превышает 6.4 процентов палинома. Разнообразные таксоны трав представлены единичными пыльцевыми зёрнами. Пыльца трав, выявленная и в ловушках, и в почвенных пробах, относится к семи семействам: Сложноцветных (Asteraceae), Зонтичных (Apiaceae), Губоцветных (Lamiaceae), Крестоцветных (Brassicaceae), Гвоздичных (Caryophyllaceae), Осоковых (Cyperaceae), Мальвовых (Malvaceae). Пыльца травянистых представителей пяти семейств: Колокольчиковых (Campanulaceae), Гераниевых (Geraniaceae), Бобовых (Fabaceae), Маковых (Papaveraceae), Кипрейных (Onagraceae) встречается только в спектрах пыльцевого дождя. Как в ловушках, так и в почвенных пробах некоторые пыльцевые зёрна не удаётся точно определить: зёрна мятые, плохой сохранности. Их количество обычно колеблется в пределах от 0.6 до трёх процентов, но очень редко достигает 11-15 процентов.
Картина спектров ловушек существенно отличаются от спектров почвенных проб содержанием спор высших растений – в трёх ловушках споры вообще не обнаружены, а в других их количество не превышает 1.33 процента.
Полученная картина имеет некоторые сходства с результатами Nosova et al. (2020). В исследованных нами спектрах тоже преобладает пыльца древесной растительности: выражены следующие доминанты Pinus, Betula, и субдоминанты Picea, Alnus. Пыльца травянистой растительности представлена в спектрах скудно, таким образом, эта экологическая группа растений «выпадает» из общей картины растительного покрова. В таёжных спорово-пыльцевых спектрах из Полистовского государственного природного заповедника также доминирующее положение занимают Pinus, Picea, Betula, Alnus (Nosova et al., 2019). Похожие тренды отмечены и в спектрах Центрально-Лесного государственного природного биосферного заповедника (Nosova et al., 2012). Изученные нами спорово-пыльцевые спектры из ловушек, в целом, соответствуют воздушным спектрам города Петрозаводска, представленным в работах Елькиной и Марковской (2007) и Елькиной (2018). Таким образом, изученные в этом исследовании спорово-пыльцевые спектры соответствуют и имеют схожие особенности с другими опубликованными спорово-пыльцевыми спектрами таёжной зоны.
Во всех изученных палиномах господствует пыльца древесных растений в пределах 58 – 98 процентов, главным образом, за счёт пыльцы хвойных, за исключением спектров из двух ловушек, в которых выявлено преимущество пыльцы лиственных пород. В большинстве образцов доминирующим таксоном является Pinus, в одном образце – Picea, а в ещё четырёх - Betula. Травянистые растения в спектрах представлены незначительно (0.3 – 7.5 процентов).
В результате исследования в спорово-пыльцевых спектрах выявлены пыльцевые зёрна следующих древесных интродуцентов: из голосеменных – Pseudotsuga menziesii, из покрытосеменных – представителей из родов Ulmus, Quercus, Juglans, Corylus, Syringa, Fraxinus, Berberis, Morus, а также представителей произрастающих на северной границе своих ареалов распространения из родов Acer и Tilia. В палиномах обычно обнаруживаются зёрна двух и более интродуцентов, но концентрация их не велика, не более двух процентов.
Пыльца орехов (Juglans) была найдена в небольшом количестве только в спектрах, расположенных относительно близко (до 1 км) от деревьев, их продуцирующих. Таким образом, пыльцу орехов (Juglans), с осторожностью, можно использовать в качестве индикатора локального произрастания орехов (Juglans).
Наличие пыльцы древесных интродуцентов обычно низкое как в ловушках, так и в почвенных пробах. На содержание пыльцы в пыльцевом дожде больше влияет расположение ловушки на дереве (интродуценте) или в непосредственной близи от него. В этих случаях содержание пыльцы растения-интродуцента может достигать 33 процентов. Однако и в этих спектрах пыльца древесных интродуцентов не господствует, доминантами остаются сосны (Pinus) и берёзы (Betula). По предварительным данным влияние одиночных экзотов распространяется не более чем на 10 - 15 метров – пыльца этих растений-интродуцентов была встречена в воздушных и почвенных спектрах не далее этого расстояния.
Палиномы отражают в основном региональную флору, локальная флора влияет на спектры при сборах на незначительном расстоянии от местных или интродуцируемых растений. Спектры из Ботанического сада и из города Петрозаводска не отличаются от типичных спектров средней тайги.
Авторы выражают благодарность и признательность всему коллективу работников Ботанического сада Петрозаводского Государственного Университета за помощь, организацию и консультации по проведению работ на территории Ботанического сада Петрозаводска.
Работа выполнена в БИН РАН (Санкт-Петербург) в рамках государственного задания по теме № 124020100138-4.
References
Elkina N. A. Pollen calendar of allergenic plants in Petrozavodsk // Scientific notes of the Petrozavodsk State University. General biology.No. 8 (177). Petrozavodsk, 2018. P. 78—82. DOI: 10.15393/uchz.art.2018.255
Elkina N. A., Markovskaya E. F. Palynological investigation of the air in the town in the taiga zone // Ecology. Experimental genetics and physiology. Works of the Karelian Scientific Center of the RAS. Vypusk 11. Petrozavodsk, 2007. P. 20—27.
Erdtman G. An Introduction to Pollen Analysis. Waltham: Chronica Botanica Company, 1943. 239 p.
Giesecke T., Fontana S. L., van der Knaap W. O., Pardoe H. S., Pidek I. A. From early pollen trapping experiments to the Pollen Monitoring Programme // Veget. Hist. Archaeobot. 19 (4). 2010. P. 247—258.
Golovko V. V. Study of the atmospheric aerosol pollen component in south of Western Siberia: Avtoref. dip. ... kand. biol. nauk. Novosibirsk, 2001. 127 p.
Gritchuk V. P., Zaklinskaya E. D. Analysis of fossil pollen and spores and its implementation in palaeogeography. M.: "Geografgiz", 1948. 223 p.
Gubankova S. G. Aeropalynology in Moscow: Avtoref. disp. … kand. biol. nauk. M., 1981. 18 p.
Hicks S., Hyvärinen H. Pollen infux values measured in different sedimentary environments and their palaeoecological implications // Grana 38. 1999. P. 228—242. https://doi.org/10.1080/00173 1300750044618
Huusko A., Hicks S. Conifer pollen abundance provides a proxy for summer temperature: Evidence from the latitudinal forest limit in Finland // J. Quat Sci. 24. 2009. P. 522—528. https://doi. org/10.1002/jqs.1250
Juggins S. C2 Version 1.5 User guide. Software for Ecological and Palaeoecological Data Analysis and Visualisation. Newcastle-upon-Tyne: Newcastle University, 2007. 73 p.
Kozlowski G., Bétrisey S., Song Y. Wingnuts (Pterocarya) and walnut family. Relict trees: linking the past, present and future. Fribourg: Natural History Museum, 2018. 128 p.
Kupriyanova L. A. Angiosperms pollen from the flora of the European part of the USSR. T. 2. L.: Nauka, 1978. 184 p.
Kupriyanova L. A. Pollen and spores of plants from the flora of the European part of the USSR. V 3 tomakh. T. 1. L.: Nauka, 1972. 171 p.
Lisitsyna O. V., Smirnov N., Aleynikov A. A. Modern pollen data from pristine taiga forest of Pechora–Ilych state nature biosphere reserve (Komi republic, Russia): first results // Ecol. Quest. 2017. 26. P. 53—55. https://dx.doi.org/10.12775/EQ.2017.016
Morozova O. V., Meshkova R. Ya. Analysis of results of aeropalynology monitoring and uptake of pollinosis patients in Smolensk region // Russian Allergology Journal. No. 4. M., 2006. P. 11—16.
Nikolskaya L. G., Fedoseev G. B. Palynological air characteristics and peculiarities of pollinosis in Leningrad // Immunologia. No. 3. M., 1987. P. 76—77.
Nosova M. B. Pollen from Holocene sequences as a source of the information about anthropogenic influence on vegetation in prehistoric period, the example from Central Forest Reserve // Bulletin of Moscow Society of Naturalists. Biological Series. 114. M, 2009. C. 30—36.
Nosova M. B. Researches of modern pollen spectra: techniques, approaches, modern areas // Botanicheskiy zhurnal. T. 105. No. 12. SPb, 2020. P. 1147—1168.
Nosova M. B., Lisitsyna O. V., Volkova O. A., Severova E. E. Variations in pollen deposition of the main taxa forming the land cover along a NW–SE transect in European Russia: results of a ten year Tauber trap monitoring period // Veget. Hist. Archaeobot. 2020. 29. P. 699—716 (2020). https://doi.org/10.1007/s00334-020-00775-1
Nosova M. B., Novenko E. Yu., Severova E. E., Volkova O. A. Vegetation and climate changes within and around the Polistovo-Lovatskaya mire system (Pskov Oblast, north-western Russia) during the past 10,500 years // Veget. Hist. Archaeobot. 2019. 28. P. 123–140. https://doi.org/10.1007/s00334-018-0693-8
Nosova M. B., Severova E. E., Volkova O. A., Kosenko J. V. Representation of Picea pollen in modern and surface samples from Central European Russia // Veget. Hist. Archaeobot. 2015. 24. P. 319—330. https://doi. org/10.1007/s00334-014-0480-0
Nosova M. B., Volkova O. A., Severova E. E. Pollen-climate relationships in broadleaved-coniferous forest zone (Central Russia) // Allergol Immunol 9. 2012. P. 179. https://doi.org/10.12775/EQ.2017.009
Novenko E. Yu., Nosova M. B., Krasnorutskaya K. V. Features of surface pollen spectra of the southern taiga on the East European Plain // News of Tula State University. Nat. Sci. 2. Tula, 2011. P. 345—354.
Petrozavodsk State University Botanic Garden; URL: http://hortus.karelia.ru/ (data: 25.01.2023).
Posevina Yu. M. Palynoecological monitoring of Ryazan city air. Avtoref. diss…. kand. biol. nauk. M., 2011. 24 p.
Severova E. E., Volkova O. A. Variations and trends of Betula pollen seasons in Moscow (Russia) in relation to meteorological parameters // Aerobiologia. 2017. 33. P. 253—264. https://doi.org/10.1007/s10453-016-9460-4
Sladkov A. N. Introduction to the pollen analysis. M.: Nauka, 1967. 275 p.
Tauber H. A static non-overload pollen collector // New Phytol. 1974. 73. P. 359—369.
