

Введение
Среди многочисленных механизмов компенсации потери воды сосудистыми растениями при транспирации и адаптации к аридным условиям, конденсация влаги на поверхности растений при достижении точки росы всегда рассматривалась только как следствие понижения температуры воздуха и поверхности почвы, обычно в ночное время.
Однако давно известно, что температура поверхности растений в большинстве случаев ниже температуры воздуха и температуры нетранспирирующих листьев (Lange, Lange, 1963). Наблюдаемое снижение температуры поверхности растений (листьев) связывается, прежде всего, с транспирацией (Gates, 1968).
Данная работа основана на предположении, что растения способны активно конденсировать воду (Прохоров, 2013) на своей поверхности более продолжительное время и более эффективно, чем почва, за счет снижения температуры поверхности ниже точки росы и увеличения доступной для конденсации поверхности.
Объекты и методы исследований
Нами для одновременного измерения температуры поверхности растений (TL) и почвы (TS), температуры (TA) и относительной влажности (HA) воздуха использовался термодетектор (пирометр) Bosch PTD 1 с функцией расчета точки росы (TD), подробные характеристики которого приведены здесь (Прохоров, 2015).
В качестве возможных конденсаторов атмосферной влаги особый интерес представляли почвопокровные растения из родов Liriope Lour. и Ophiopogon Ker.-Gawl., в отношении которых в Субтропическом ботаническом саду Кубани (СБСК) проводились исследования по их пригодности для создания вечнозеленых газонных покрытий в теневых местах урбанизированных экосистем.
Результаты и обсуждение
Ранее, опытным путём, была установлена аномально высокая засухоустойчивость представителей родов Liriope Lour. и Ophiopogon Ker.-Gawl. (Коннов, 2014; Коннов, Карпун, 2014), нетипичная для растений влажного муссонного климата Восточной Азии (табл. 1).
Таблица 1. Динамика изменения оводнённости листьев представителей родов Liriope и Ophiopogon в условиях искусственной засухи. Сочи, СБСК, 07–08. 2014 г.
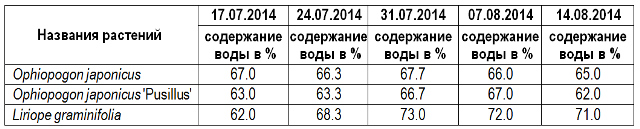
Как видно из приведённой таблицы, содержание воды в листьях опытных растений после месячной искусственной засухи не только практически не снизилось, но у Liriope graminifolia даже возросло. Такой результат, казалось бы, должен противоречить общепринятой точке зрения на поведение растений в условиях постоянного дефицита воды.
Именно это обстоятельство побудило нас провести замеры температуры поверхности ряда растений из рассматриваемых родов (табл. 2).
Полученные результаты показывают, что температура поверхности листьев была значительно ниже не только температуры воздуха, но и, за исключением Ophiopogon jaburan 'Vittatus', ниже точки росы. И, соответственно, происходит конденсация атмосферной влаги на поверхности листьев.
Попытка провести анализ зависимости степени снижения температуры поверхности листьев от биологических особенностей растений не дала желаемых результатов. Так, наиболее низкую температуру имели и Ophiopogon umbraticola, с его узкими листьями, шириной 2 мм, и Ophiopogon planiscapus, у которого ширина листьев превышает 10 мм. Вид из южного Китая, Ophiopogon sarmentosus, имел температуру поверхности листьев 15.8° С, а у самого северного вида рода, Ophiopogon japonicus, она была 13.3° С. Существенно отличались по температуре листьев растения Ophiopogon planiscapus 'Nigrescens' семенного и вегетативного происхождения, 16.2° С и 13.2° С, соответственно. Однако, в целом, листья представителей рода Liriope были заметно «теплее», нежели листья представителей рода Ophiopogon.
Таблица 2. Температура поверхности (TL) контейнерных растений Liriope и Ophiopogon и микроклиматические параметры (TA, HA, TD). Сочи, СБСК, 10.06.2015 г. 12:00, тень
| Наименование растений | TL,°C | TA,°C | HA,% | TD,°C |
| Liriope exiliflora | 16.7 | 24.3 | 62 | 17.2 |
| Liriope muscari 'Variegata' | 16.4 | 24.3 | 62 | 17.2 |
| Liriope graminifolia | 15.4 | 24.3 | 62 | 17.2 |
| Liriope koreana | 16.0 | 24.3 | 62 | 17.2 |
| Liriope muscari 'Royal Purple' | 16.0 | 24.3 | 62 | 17.2 |
| Liriope spicata 'Variegata' | 16.0 | 24.3 | 62 | 17.2 |
| Liriope minor | 15.5 | 24.3 | 62 | 17.2 |
| Liriope zhejiangensis | 15.0 | 24.3 | 62 | 17.2 |
| Liriope muscari | 13.5 | 24.3 | 62 | 17.2 |
| Ophiopogon jaburan 'Vittatus' | 17.8 | 24.3 | 62 | 17.2 |
| Ophiopogon jaburan | 16.7 | 24.3 | 62 | 17.2 |
| Ophiopogon planiscapus 'Nigrescens' , сем. | 16.2 | 24.3 | 62 | 17.2 |
| Ophiopogon chingii | 16.0 | 24.3 | 62 | 17.2 |
| Ophiopogon sarmentosus | 15.8 | 24.3 | 62 | 17.2 |
| Ophiopogon intermedius | 15.6 | 24.3 | 62 | 17.2 |
| Ophiopogon japonicus 'Pusillus', сем. | 15.4 | 24.3 | 62 | 17.2 |
| Ophiopogon japonicus for. media | 15.1 | 24.3 | 62 | 17.2 |
| Ophiopogon japonicus 'Pusillus', дел. | 15.1 | 24.3 | 62 | 17.2 |
| Ophiopogon stenophyllus | 13.5 | 24.3 | 62 | 17.2 |
| Ophiopogon japonicus | 13.3 | 24.3 | 62 | 17.2 |
| Ophiopogon planiscapus 'Nigrescens', дел. | 13.2 | 24.3 | 62 | 17.2 |
| Ophiopogon umbraticola | 12.9 | 24.3 | 62 | 17.2 |
| Ophiopogon planiscapus | 12.7 | 24.3 | 62 | 17.2 |
| Примечания: сем. – из семян; дел. – деление растения. | ||||
Учитывая незначительный объём надземной части коллекционных растений в контейнерах, и, как следствие этого, существенное влияние температуры почвы на температуру поверхности листьев, дальнейшие исследования проводились на густо задернённых опытных делянках, заложенных в 2010 году. Соответствующие замеры на трёх таксонах, наиболее массово и давно культивируемых в регионе, проводились с интервалами в 4 часа (рис. 1–3).
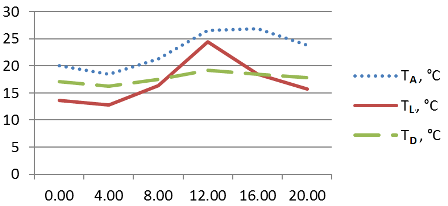
Рис. 1. Суточный ход температурных параметров делянки с Ophiopogon japonicus. Сочи, СБСК, 10–11.06.2015 г.
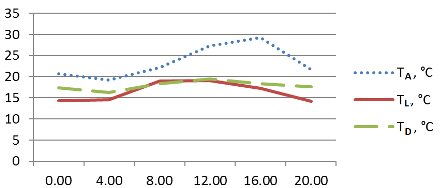
Рис. 2. Суточный ход температурных параметров делянки с Ophiopogon japonicus 'Pusillus'. Сочи, СБСК, 10–11.06.2015 г.
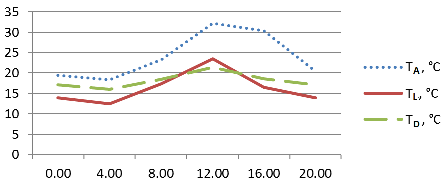
Рис. 3. Суточный ход температурных параметров делянки с Liriope graminifolia. Сочи, СБСК, 10–11. 06. 2015 г.
Для всех выбранных таксонов температура поверхности листьев была существенно ниже температуры воздуха. Максимальный разрыв составлял от 12,7° C (Liriope graminifolia) до 10,9° C (Ophiopogon japonicus), тогда как минимальный разрыв составил 3,6° C (Ophiopogon japonicus 'Pusillus' ). Причём максимальные температуры воздуха и поверхности листьев отмечались у Ophiopogon japonicus и Liriope graminifolia в 12:00 (рис. 1, 3), тогда как у карликовой формы первого виды максимальная температура воздуха была зафиксирована в 16:00, а максимальная температура листьев – утром, в 8:00 (рис. 2). Температура поверхности листьев повышалась выше показателя точки росы ненадолго в дневные часы – более выражено у Ophiopogon japonicus и незначительно у его садовой формы (рис. 1–2). Следует отметить, что потенциальная способность к конденсации влаги из атмосферного воздуха на поверхности опытных растений отмечалась, в среднем, 19 часов и была наиболее продолжительной у Liriope graminifolia – 20 часов, с 15:00 10.06 до 11:00 11.06.
Результаты проведённых измерений побудили нас проанализировать характер потери влаги этими таксонами в течение светлого и тёмного времени суток, для чего был проведён соответствующий опыт. Двулетние растения в контейнерах объёмом 1,5 л и такие же контейнеры, заполненные аналогичной почвосмесью, были выставлены на площадке под открытым небом и под густой кроной дерева (Ficus carica). Погода была солнечной; средняя ночная температура составляла 22° C, средняя дневная – 26° C. Взвешивания производились на электронных весах в 6:00 и 18:00. Анализ полученных данных по фактической потере влаги в граммах, в пересчёте на килограмм общего веса, приведён в таблицах 3 и 4.
Таблица 3. Анализ потери влаги контейнерами с растениями. Сочи, СБСК, 11 – 13. 06.2015 г.

Таблица 4. Анализ потери влаги контейнерами с растениями. Сочи, СБСК, 11–13. 06.2015 г.
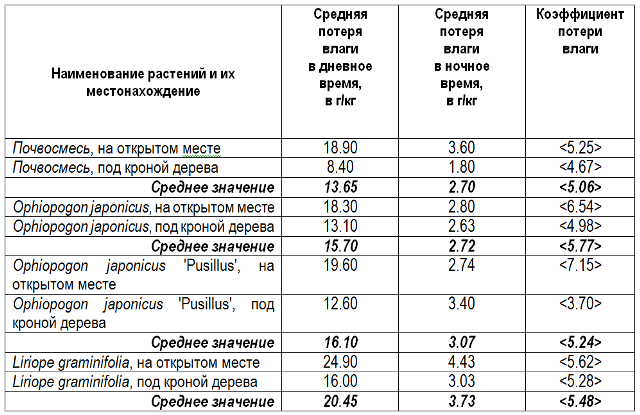
Во всех случаях разница в потере влаги контейнерами с растениями по отношению к контейнерам без растений не превышала 2 г на кг, будучи наиболее низкой у Ophiopogon japonicus и наиболее высокой у Liriope graminifolia. И это притом, что поверхность, с которой испаряется влага, у контейнеров с растениями была значительно больше, соответственно и влаги должно было испариться значительно больше, нежели было зафиксировано. Такое несоответствие можно объяснить только тем, что часть испарившейся влаги компенсировалась за счет конденсации атмосферной влаги на листьях, когда их температура была ниже точки росы. При этом не обязательно, чтобы влага вначале была поглощена листьями, а уже затем испарилась через устьица.
В подтверждение этого может свидетельствовать и то, что в дневное время интенсивность испарения возрастала более, чем в пять раз (табл. 4), тогда как средняя дневная температура была больше средней ночной всего лишь на 4o C. Но в дневное время температура поверхности листьев была либо выше точки росы, либо незначительно ниже и, соответственно, количество влаги, конденсировавшейся на поверхности листьев, было меньше, чем в ночное время.
Заключение
Результаты проведённых исследований позволяют под новым углом зрения взглянуть на терморегуляцию у растений. Существующие представления, что более низкая температура поверхности листьев растений есть следствие интенсивного испарения влаги и способствует, преимущественно, предохранению растений от дневного перегрева (Curtis, 1936), не вполне корректно объясняют это явление. По нашему мнению, терморегуляция растений многофункциональна и одним из ее предназначений может являться самоорошение путем конденсации атмосферной влаги.
Нельзя исключить адаптационное значение конденсации атмосферной влаги на поверхности растений, охлажденной ниже точки росы. Отсюда вытекает значимость понимания данного явления при интродукционных испытаниях растений в несвойственных ему климатических условиях.
Литература
Коннов Н. А. Офиопогон и Лириопа – перспективные газонные растения // Цветоводство . 2014. № 6. С. 12—14.
Коннов Н. А., Карпун Ю. Н. Итоги и перспективы интродукции представителей рода Ophiopogon Ker Gawler на Черноморское побережье России // Труды Кубанского аграрного университета. 2014. Вып. 3(48). С. 57—61.
Прохоров А. А. Активная конденсация воды растениями // Принципы экологии. ПетрГУ. 2013. № 3. С. 58—61. DOI: 10.15393/j1.art.2013.2921.
Прохоров А. А. Точка росы - неизученный фактор в экологии, физиологии и интродукции растений // Hortus bot. 2015. Т. 10. DOI: 10.15393/j4.art.2015.2801.
Curtis O. F. Leaf temperatures and the cooling of leaves by radiation // Plant Physiol. 1936. Apr; 11(2). P. 343—364.
Gates D. M. Transpiration and Leaf Temperature // Annual Review of Plant Physiology. 1968. Vol. 19. P. 211—238.
Lange O. L., Lange R. Untersuchungen uber Blattemperaturen, Transpiration und Hitzeresistenz an Pflanzen mediterraner Standorte (Costabrava, Spanien). // Flora. 1963. 153. S. 387—425.
