

Введение
Глобальные изменения климата, а также особенности проявления этих процессов на региональном уровне — наиболее обсуждаемая проблема последних десятилетий. Понимание взаимосвязей климата и фенологии растений на природных и урбанизированных территориях вызывает широкий интерес в научных кругах (CaraDonna et al., 2014; Bjorkman et al., 2015; Минин и др., 2018). Актуальна как первичная информация о сезонном развитии растений, так и разработка методов современной обработки данных (Cleland et al., 2007; Wolkovich, Cleland, 2011; Stucky et al., 2018). Эта тема имеет важнейшее значение для ботанических садов, где традиционно исследуется влияние местных климатических условий на рост и развитие растений в условиях культуры. Ряд садов имеют неоценимые по своему научному значению многолетние базы данных фенологических и метеорологических наблюдений, позволяющие отслеживать тенденции изменений климата и реакции различных видов растений (Булыгин, Довгулевич, 1974; Фирсов, 2014). Актуальность исследований ботанических садов в области мониторинга изменения климата, смягчения его последствий для природных комплексов и человека, планирование мер для поддержания коллекций растений рассматриваются на конгрессах ботанических садов и тематических конференциях (Schulman, Lehvävirta, 2011; Современное состояние фенологии …, 2015; Летопись природы России …, 2018).
В Ботаническом саду ПетрГУ фенологические исследования проводились с 1960-х годов, но, к сожалению, не на постоянной основе. На сегодняшний день мы имеем разрозненные данные по отдельным группам таксонов, собранные в течение различных временных периодов. Особенности роста и развития древесных растений в коллекциях Ботанического сада достаточно полно представлены в публикациях И. Т. Кищенко с соавторами (2000, 2008, 2015 и др.), А. С. Лантратовой с соавторами (1962, 1969, 1971, 1979 и др.). Результаты фенологических наблюдений за декоративными травянистыми растениями практически не были отражены в печати. Большее внимание в предшествующие годы уделялось исследованиям сезонного развития однолетних, многолетних пищевых, кормовых и лекарственных травянистых культур (Штанько, 1975; Штанько, Поташева, 1982; Штанько А. В., Штанько С. А., 1993 и др.).
В настоящее время проводится работа по восстановлению исторических данных и их обобщению. Организованы систематические фенологические наблюдения за растениями коллекционных фондов Ботанического сада. В данной работе приводятся первые результаты исследований сезонного развития травянистых растений экспозиции «Теневой сад» в связи с климатическими параметрами за период 2016-2018 гг.
Объекты и методы исследований
Ботанический сад ПетрГУ расположен в подзоне средней тайги, в четвертой зоне зимостойкости. Общая площадь сада 367 га, коллекции и экспозиции культурных растений располагаются на 37 га, остальную часть занимает природная территория. Коллекции сада насчитывают более 2600 таксонов, в экспозиции «Теневой сад» выращиваются 250 видов, подвидов и сортов травянистых растений. Экспозиция располагается под пологом сосен, которые создают разреженную тень в течение всего вегетационного сезона.
В данном исследовании представлены результаты наблюдений за 44 таксонами растений; среди них 39 видов, один гибридный вид, принадлежащие 30 родам, 17 семействам. Наблюдались сорта (5), в том числе гибридного происхождения, у ряда видов - подвиды (3). По географическому распространению большинство видов являются евразиатскими (15), также присутствуют азиатские (10), европейские (9) и американские (5). Десять видов являются аборигенными, семь из них (Hepatica nobilis, Anemone ranunculoides, Ficaria verna, Campanula latifolia, Carex muricata, Galium odoratum, Thalictrum aquilegiifolium) включены в Красную книгу Карелии (2007). По жизненной форме все исследуемые растения - многолетние поликарпики, на период наблюдений находились в генеративном онтогенетическом состоянии.
Согласно традиционной методике фенологических наблюдений (Методика …, 1975) фиксировали даты следующих фенофаз: начало весеннего отрастания (или начало вегетации для зимнезеленых растений), начало бутонизации, начало цветения, конец цветения, завязывание плодов, созревание плодов, раскрывание или опадение плодов, осыпание семян, засыхание или расцвечивание листьев, полное увядание. Феноритмотипы растений рассматривались согласно И. В. Борисовой (1965) и Р. А. Карписоновой (1985).
В работе использовали данные метеостанции Davis Vantage Pro 2 Plus Ботанического сада ПетрГУ и Сулажгорской метеостанции (г. Петрозаводск, номер в ВМО (WMO ID) 22820), расположенной в 3 км к юго-западу от Ботанического сада. В Ботаническом саду измерялись среднесуточная температура воздуха о С, рассчитанная как среднее 48 измерений через каждые полчаса. Остальные метеоданные, включая средние значения температуры и осадков за последние 10 лет, брали из архива Сулажгорской метеостанции.
Результаты и обсуждение
Сравнение среднемесячных значений температуры и осадков показало довольно значительное их варьирование в течение 2016-2018 гг. наблюдений (табл. 1). Более теплыми, по сравнению с многолетними данными, в 2016 году были весенние месяцы и первая половина лета. Количество осадков в этом году превышало средние многолетние значения в 2-2,5 раза в июле и августе. В 2018 году весна была ранней и теплой, без возвратных заморозков. Более теплым был практически весь вегетационный сезон. Приход солнечной радиации был в несколько раз выше в 2018 г. по сравнению с 2017 годом. Количество осадков в 2018 г. было близким к норме за исключением резкого снижения этого показателя в мае и превышения - в апреле и августе. 2017 год характеризовался поздним наступлением тепла, более низкими температурами за период с апреля по июль. Количество осадков в этот год в апреле, мае, сентябре превышало средние многолетние данные примерно в три раза.
Таблица 1. Температура воздуха и осадки в течение вегетационных периодов 2016–2018 гг.
| Год | апрель | май | июнь | июль | август | сентябрь | октябрь |
| среднесуточные температуры воздуха, о С | |||||||
| 2016* | -1,7/10,3 3,5 |
6,2/17,2 12,7 |
6,7/20,1 14,2 |
14/22,9 18,5 |
8,4/19,1 15,3 |
6,4/15,5 10 |
-0,8/9,3 2,8 |
| 2017 | -6,0 / 6,5 0,2 |
-0,4 / 11,4 5,2 |
4,2 / 18,3 11,9 |
11,0 / 19,4 15,7 |
9,8 / 19,4 16,1 |
6,8 / 15,8 10,1 |
-1,8 / 8,2 3,8 |
| 2018 | -3,4 / 8,7 3,4 |
2,6 / 19,9 12,5 |
7,1 / 20,5 13,8 |
11,7 / 23,6 18,9 |
11,3 / 25,3 17,1 |
3,2 / 18,8 12,0 |
-3,8/13,8 5,3 |
| Средние многолетние* | 2,6 | 9,8 | 13,7 | 17,6 | 15,4 | 10,4 | 3,8 |
| месячные суммы осадков, мм | |||||||
| 2016* | 101,2 | 89,9 | 135,6 | 254,1 | 398 | 98,1 | 23,9 |
| 2017* | 113,3 | 310,5 | 136,1 | 149,8 | 122,9 | 276,8 | 90,7 |
| 2018* | 128,1 | 34,1 | 74 | 114,8 | 210,3 | 100,1 | 84,9 |
| Средние многолетние* | 47,2 | 93,4 | 97,5 | 120,4 | 141,0 | 100,6 | 71,4 |
* Среднемесячные значения температуры за 2016 г., суммы осадков за 2016-2018 гг., средние многолетние значения температуры рассчитаны по данным архива метеонаблюдений https://rp5.ru, полученным на метеостанции в Петрозаводске (р-н Сулажгора), номер в ВМО (WMO ID) 22820.
Значительное различие погодных условий в течение трех лет наблюдений предполагает получение более полной информации о варьировании сроков наступления фенофаз исследуемых растений.
Начало вегетации
Сход снега в «Теневом саду» происходит в последней декаде апреля. В эти сроки начинается вегетация видов, зимующих с зелеными листьями: Bergenia crassifolia (рис. 1), Carex muricata, Epimedium × rubrum, Heuchera cylindrica, H. sanguinea, Primula auricula, P. denticulata, P. juliae, Pulmonaria rubra, P. saccharata, Saxifraga umbrosa, Tiarella wherryi (табл. 2-5). Для условий Западной Сибири показано, что при благоприятных условиях перезимовки старые листья зимнезеленых поликарпиков фотосинтезируют не только раньше по срокам, но и интенсивнее молодых (Фомина, 2012). У некоторых видов растений (Geranium phaeum, G. sanguineum) листья увядают в течение зимы или сохраняются в отдельные годы (Luzula nivea, Epimedium × rubrum). Развертывание новых листьев у большинства образцов этой феноритмологической группы в теплые годы в Карелии происходит в первой декаде мая; у Epimedium × rubrum новые побеги отрастали во второй-третьей декаде мая, после усыхания зимующих листьев.
Ранняя вегетация (конец апреля - первые числа мая) характерна также для Ficaria verna, Hemerocallis middendorffii, Hepatica nobilis, Primula veris, которые способны фотосинтезировать уже при низких положительных температурах. Так, в течение трех лет наблюдений, к 1 мая сумма температур выше 0° (расчет проводили с 15 апреля) составляла 18-67°, в те же сроки сумма температур выше +5° — 5-50°.
В 2017 году наблюдался поздний сход снега и затянувшаяся холодная весна, в связи с чем у ряда растений был отмечен сдвиг сроков отрастания на 10-20 дней. Таким образом, можно предполагать их большую требовательность к теплу на начальном этапе роста. К таким видам относятся: Anemone ranunculoides, Carex muricata, Epimedium × rubrum, Geranium phaeum, Heuchera cylindrica, H. sanguinea, Iris sibirica и его сорт 'Purple mere', Saxifraga umbrosa, Thalictrum aquilegiifolium. По результатам трех лет исследований, к моменту начала вегетации этих растений, сумма температур выше 0° составляла 50-140°, выше 5° — 26-96°. Весеннее отрастание Geranium sanguineum, Iris versicolor 'Kermensina', Luzula nivea, Tiarella wherryi происходило в пределах такой же суммы температур, но связь с изменением температурного режима в 2017 г. не наблюдалась.
Во второй декаде мая в 2016, 2018 гг. с запозданием на третью декаду в 2017 году вегетация начиналась у следующих видов: Aruncus dioicus, Podophyllum peltatum, Rodgersia podophylla, Thalictrum simplex, Veronicastrum sibiricum. Еще более поздние сроки начала вегетации (первые числа июня) в 2017 году отмечали для Clematis recta, Convallaria majalis, Digitalis lutea, Galium odoratum. Сумма активных температур выше 5° в этот период составляла 82-196°. Интересной особенностью является практически синхронное пробуждение и развитие растений этой группы в весенний период. Необходимым условием являлся переход среднесуточных температур через рубеж 15° в 2016 и 2018 годах и 10° – в 2017 г.
Самые поздние сроки отрастания – в конце мая – первых числах июня – наблюдались у видов и сортов рода Hosta Tratt. В 2017 г. у этих растений был существенный сдвиг (около 3 недель) в наступлении фенофазы. Начало вегетации происходило после установления среднесуточных температур выше 13-15°, что соответствует данным, полученным в Центральном сибирском ботаническом саду СО РАН (Седельникова, 2012). Сумма активных температур выше 5° в период отрастания в течение трех лет наблюдений варьировала в широких пределах 142-480°.

Рис. 1. Bergenia crassifolia в апреле.

Рис. 2. Lamprocapnos spectabilis. Начало вегетации.
Начало цветения
Все исследуемые таксоны ежегодно цветут в условиях Карелии, большинство относится к группе декоративноцветущих растений, что открывает широкие возможности их использования в озеленении.

Рис. 3. Цветение Pulmonaria rubra.

Рис. 4. Цветение Primula denticulata.

Рис. 5. Бутонизация Podophyllum peltatum.
По срокам цветения к ранневесеннему феноритмотипу в условиях местного климата можно отнести Hepatica nobilis (распускание бутонов происходит практически сразу после схода снега в конце апреля - начале мая). Во второй половине мая цветут поздневесенние Anemone ranunculoides, Ficaria verna, Epimedium × rubrum, Primula auricula, P. denticulata (рис. 4), P. juliae 'Gartengluck', P. veris, Pulmonaria rubra (рис. 3), P. saccharata.
На конец мая - начало июня приходится цветение раннелетних растений: Galium odoratum, Tiarella wherryi. Самую большую группу составляют среднелетние растения, цветущие с конца июня до середины июля: Aruncus dioicus, Campanula latifolia, Carex muricata, Clematis recta, Convallaria majalis, Geranium phaeum, G. sanguineum, Iris sibirica, I. sibirica 'Purple mere', I. versicolor 'Kermensina', Hemerocallis middendorffii, Luzula nivea, Podophyllum peltatum (рис. 5), Thalictrum aquilegiifolium, Thalictrum simplex.
К позднелетним по срокам цветения (конец июля - середина августа) относятся Digitalis lutea, Veronicastrum sibiricum.
Наиболее варьируют по годам (около месяца) даты начала цветения Clematis recta, Carex muricata, Hosta sieboldiana var. elegans, Thalictrum simplex, Tiarella wherryi, Veronicastrum sibiricum, 3-4 недели – Aruncus dioicus, Galium odoratum, Geranium sanguineum, Hemerocallis middendorffii, Heuchera cylindrica, Iris sibirica, Ligularia sibirica, Thalictrum aquilegiifolium, около 3-х недель – Campanula latifolia, Convallaria majalis, Digitalis lutea, Ficaria verna, Heuchera sanguinea, Hosta sieboldiana var. aureomarginata, Iris sibirica 'Purple mere', Luzula nivea, Primula denticulata, Pulmonaria rubra, 2-3 недели – Bergenia crassifolia, Epimedium × rubrum, Geranium phaeum, Hosta undulata var. albomarginata, Hosta 'Devon Green', Iris versicolor 'Kermensina', Lamprocapnos spectabilis, Podophyllum peltatum, P. veris, Pulmonaria saccharata, Saxifraga umbrosa, около 10 дней - 2 недель – Anemone ranunculoides, Hepatica nobilis, Primula juliae 'Gartengluck', Rodgersia podophylla, Scilla luciliae, Hosta 'Canadian blue', Scilla siberica, около недели – Aconitum napellus.
Практически не варьируют по годам даты начала цветения Hosta sieboldiana var. elegans.
В целом, варьирование сроков цветения не связано с принадлежностью к определенному феноритмотипу, а определяется видовыми и сортовыми особенностями.

Рис. 6. Лето в «Теневом саду».
Разница в наступлении сроков цветения может быть результатом реакции видов на погодные условия, на севере ведущим является температурный фактор. К сожалению, трехлетний период наблюдений не позволяет статистически подтвердить связь сроков бутонизации и цветения с суммой эффективных температур, но, тем не менее, наличие сезонов с контрастными погодными условиями дает возможность предполагать эту зависимость у ряда исследуемых видов.
Так, близкие значения суммы температур выше 5° отмечаются в период начала бутонизации Anemone ranunculoides (147-196°), Aruncus dioicus (382-431°), Convallaria majalis (301-379°), Dicentra spectabilis (173-243°), Digitalis lutea (899-970°), Geranium sanguineum (431-459°), Hemerocallis middendorffii (424-446°), Heuchera sanguinea (372-431°), Primula auricula (116-148°), P. veris (27-68°), Saxifraga umbrosa (302-379°), Thalictrum simplex (481-486°), для дат начала цветения Carex muricata (431-507°), Clematis recta (827-869°), Digitalis lutea (1067-1086°), Epimedium × rubrum (229-265°), Ficaria verna (173-176°), Galium odoratum (420-424°), Hepatica nobilis (27-65°), Primula denticulata (83-148°), Pulmonaria saccharata (143-197°), Tiarella wherryi (358-420°), для плодоношения Pulmonaria rubra (397-431°). Для остальных исследуемых видов растений варьирование суммы температур в пределах дат начала бутонизации и цветения имеет широкий диапазон.

Рис. 7. Geranium phaeum. Начало плодоношения.

Рис. 8. Цветы и плоды Iris versicolor 'Kermensina'.
Продолжительность цветения
Продолжительность периода цветения у исследуемых образцов коллекции «Теневой сад» варьирует в широких пределах (табл. 5, рис. 6). Менее длителен этот период у группы растений с ранним сроком цветения (Epimedium × rubrum, Ficaria verna, Hepatica nobilis, Primula veris и др.). Растянутый период цветения наблюдается у растений позднелетней группы, таких как Digitalis lutea, Thalictrum simplex, Veronicastrum sibiricum. Этот период у Clematis recta, Geranium phaeum, G. sanguineum, Tiarella wherryi увеличивается за счет волн повторного цветения, продолжающегося до осени. Длительное цветение наблюдается также у растений с постепенным развитием цветков в акропетальном направлении: Dicentra spectabilis, Ligularia sibirica, Pulmonaria rubra, P. saccharata. Подобные биологические адаптации увеличивают возможность опыления и надежность семенного размножения.
Интересной особенностью является довольно значительные изменения продолжительности цветения у некоторых образцов в более холодные и влажные годы. Это явление ранее наблюдалось Р. А. Карписоновой (1985) для некоторых декоративных растений в условиях Главного ботанического сада имени Н. В. Цицина РАН. Так в 2017 году период цветения превышал более чем на 10 дней у Primula juliae, Anemone ranunculoides, Iris versicolor 'Kermensina', Veronicastrum sibiricum, Hosta sieboldiana var. aureomarginata, более чем на 20 дней - у Digitalis lutea, Hosta sieboldiana var. elegans, Hosta 'Canadian blue'. И, наоборот, у Ligularia sibirica он был более коротким по сравнению с 2016 и 2018 годами.
Созревание семян
Успешность семенного размножения является одной из главных характеристик адаптации растений в условиях интродукции. В коллекции растений «Теневой сад» ежегодно образуют семена 35 таксонов (рис. 7, 8). Целый ряд видов имеют самосев: Aruncus dioicus, Campanula latifolia, Clematis recta, Geranium sanguineum, G. phaeum, Hepatica nobilis, Heuchera cylindrica, H. sanguinea, Lamprocapnos spectabilis, Ligularia sibirica, Primula veris, P. denticulata, Pulmonaria rubra, P. saccharata.
В течение трех лет наблюдений плоды образовывались, но семена не вызревали у Hosta 'Canadian blue', Hosta undulata var. albomarginata, H. sieboldiana var. aureomarginata, H. sieboldiana var. elegans (подтверждено экспериментами по определению полевой всхожести). Не ежегодно происходит образование плодов Podophyllum peltatum. Не наблюдалось завязывания плодов у Primula juliae 'Gartengluck' и Hosta 'Devon Green'.

Рис. 9. Geranium sanguineum. Осенняя окраска листьев.
Продолжительность вегетации
Небольшим числом представлены растения с относительно коротким периодом вегетации: эфемероиды (3) и весенне-летнезеленые (4). Период вегетации эфемероидов (Ficaria verna, Scilla siberica, Scilla luciliae) длится 35-70 дней. У всех трех видов эфемероидов в 2017 году этот период был более продолжительным на 10-20 дней. Вегетационный период у весенне-летнезеленых растений продолжается с весны до начала осени и составляет 100-140 дней.
Период со стабильными положительными температурами заканчивается в условиях Карелии во второй половине октября. В эти же сроки наблюдаются заморозки и первый снег, заканчивают вегетацию весеннее-летне-осеннезеленые и весеннее-летне-зимнезеленые растения (рис. 9). Представители первой группы преобладают в коллекции (20 таксонов), период вегетации у них продолжается 120-190 дней. Вегетация весенне-летне-зимнезеленых растений (16 таксонов) прерывается низкими температурами и устойчивым снежным покровом осенью (середина ноября) и восстанавливается весной.
Таблица 2. Даты наступления фенофаз растений весеннего цветения в экспозиции «Теневой сад»
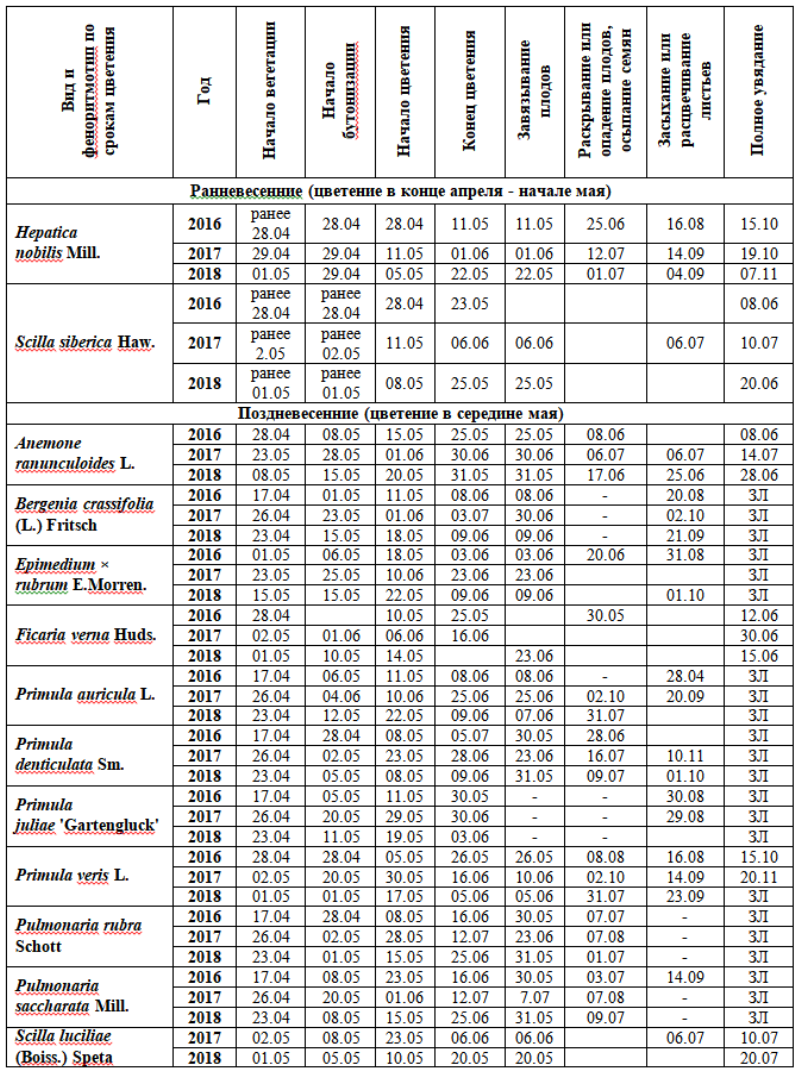
ЗЛ – зимой сохраняется надземная часть растения полностью или частично, - фенофаза отсутствовала, пустые ячейки – нет данных.
Таблица 3. Даты наступления фенофаз растений ранне- и среднелетнего цветения в экспозиции «Теневой сад»
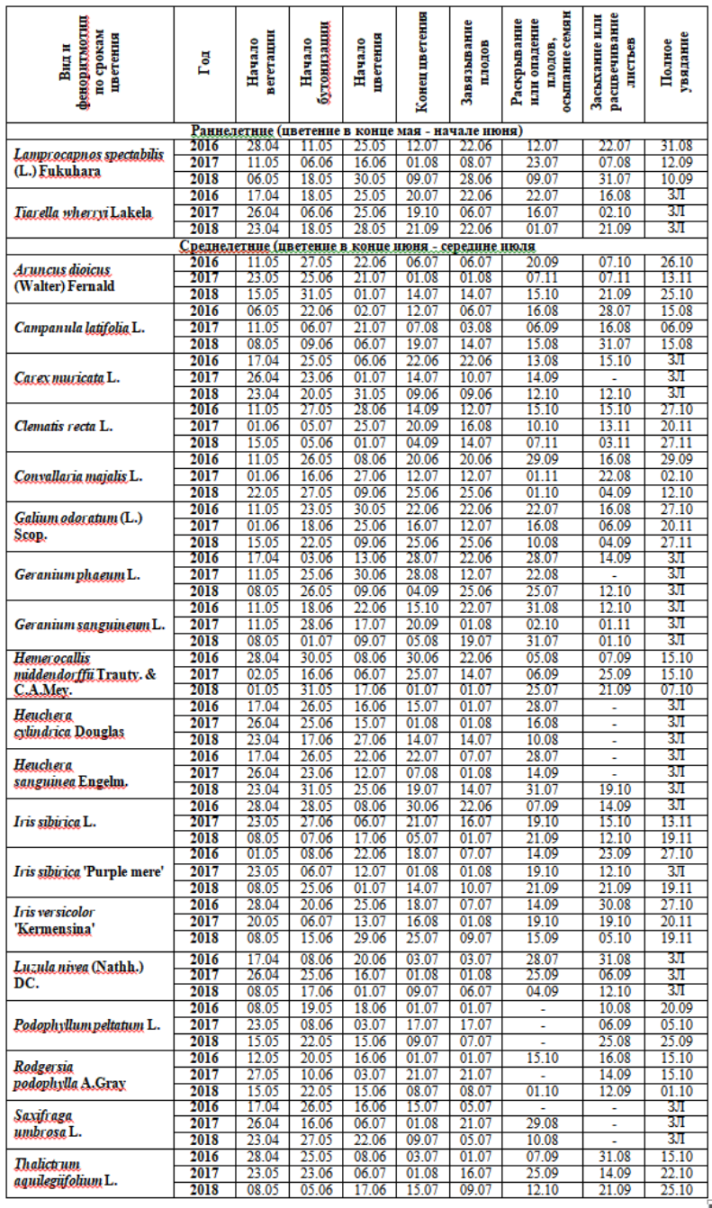
ЗЛ – зимой сохраняется надземная часть растения полностью или частично, - фенофаза отсутствовала.
Таблица 4. Даты наступления фенофаз растений позднелетнего и осеннего цветения в экспозиции «Теневой сад»

- фенофаза отсутствовала, пустые ячейки – нет данных.
Таблица 5. Феноритмотипы коллекции многолетних растений «Теневой сад» по срокам цветения и продолжительности вегетации
| Вид и феноритмотип по срокам цветения | Продолжительность цветения, дни | Феноритмотип по длительности вегетации |
| Ранневесенние (конец апреля - начало мая) | ||
| Hepatica nobilis Mill. | 14-18 | Весеннее-летне-осеннезеленый |
| Scilla siberica Haw. | 18-27 | Эфемероид |
| Поздневесенние | ||
| Anemone ranunculoides L. | 11-25 | Эфемероид |
| Bergenia crassifolia (L.) Fritsch | 23-33 | Весеннее-летне-зимнезеленый |
| Epimedium × rubrum E. Morren. | 14-19 | Весеннее-летне-зимнезеленый |
| Ficaria verna Huds. | 11-16 | Эфемероид |
| Primula auricula L. | 16-19 | Весеннее-летне-зимнезеленый |
| Primula denticulata Sm. | 33 | Весеннее-летне-зимнезеленый |
| Primula juliae 'Gartengluck' | 16-30 | Весеннее-летне-зимнезеленый |
| Primula veris L. | 18-22 | Весеннее-летне-зимнезеленый |
| Pulmonaria rubra Schott | 40-45 | Весеннее-летне-зимнезеленый |
| Pulmonaria saccharata Mill. | 25-42 | Весеннее-летне-зимнезеленый |
| Scilla luciliae (Boiss.) Speta | 11-15 | Эфемероид |
| Раннелетние (конец мая - начало июня) | ||
| Lamprocapnos spectabilis (L.) Fukuhara | 37-49 | Весенне-летнезеленый |
| Tiarella wherryi Lakela | 57-58 | Весеннее-летне-зимнезеленый |
| Среднелетние (конец июня - середина июля | ||
| Aruncus dioicus (Walter) Fernald | 12-15 | Весеннее-летне-осеннезеленый |
| Campanula latifolia L. | 11-18 | Весенне- летнезеленый |
| Carex muricata L. | 11-17 | Весеннее-летне-зимнезеленый |
| Clematis recta L. | 58-79 | Весеннее-летне-осеннезеленый |
| Convallaria majalis L. | 13-17 | Весенне- летнезеленый |
| Galium odoratum (L.) Scop. | 18-24 | Весеннее-летне-осеннезеленый |
| Geranium phaeum L. | 46-88 | Весеннее-летне-зимнезеленый |
| Geranium sanguineum L. | 66-90 | Весеннее-летне-зимнезеленый |
| Hemerocallis middendorffii Trautv. & C. A. Mey. | 14-23 | Весеннее-летне-осеннезеленый |
| Heuchera cylindrica Douglas | 18-30 | Весеннее-летне-зимнезеленый |
| Heuchera sanguinea Engelm. | 25-31 | Весеннее-летне-зимнезеленый |
| Iris sibirica L. | 16-23 | Весеннее-летне-осеннезеленый |
| Iris sibirica 'Purple mere' | 15-27 | Весеннее-летне-осеннезеленый |
| Iris versicolor 'Kermensina' | 24-35 | Весеннее-летне-осеннезеленый |
| Luzula nivea (Nathh.) DC. | 10-15 | Весеннее-летне-зимнезеленый |
| Podophyllum peltatum L. | 14-25 | Весенне- летнезеленый |
| Rodgersia podophylla A. Gray | 16-24 | Весенне- летнезеленый |
| Saxifraga umbrosa L. | 18-30 | Весеннее-летне-зимнезеленый |
| Thalictrum aquilegiifolium L. | 26-29 | Весеннее-летне-осеннезеленый |
| Позднелетние (конец июля - середина августа) | ||
| Aconitum napellus L. | 16-20 | Весеннее-летне-осеннезеленый |
| Digitalis lutea L. | 38-65 | Весеннее-летне-осеннезеленый |
| Hosta sieboldiana var. aureomarginata Makino | 17-39 | Весеннее-летне-осеннезеленый |
| Hosta sieboldiana var. elegans Hyl. | 23-56 | Весеннее-летне-осеннезеленый |
| Hosta undulata var. albomarginata F. Maek. | 22-31 | Весеннее-летне-осеннезеленый |
| Ligularia sibirica (L.) Cass. | 22-38 | Весеннее-летне-осеннезеленый |
| Thalictrum simplex L. | 30-39 | Весеннее-летне-осеннезеленый |
| Veronicastrum sibiricum (L.) Pennell. | 34-59 | Весеннее-летне-осеннезеленый |
| Осенние (сентябрь) | ||
| Hosta 'Canadian blue' | 31-65 | Весеннее-летне-осеннезеленый |
| Hosta 'Devon Green' | 40-49 | Весеннее-летне-осеннезеленый |
Выводы и заключение
Установлены продолжительность вегетации и сроки основных фенофаз для 44 таксонов травянистых растений экспозиции «Теневой сад» в условиях Карелии.
В течение трех лет наблюдений отмечены резкие отклонения среднемесячных температур по сравнению со средними многолетними данными в сторону более высоких значений в первую половину вегетационного сезона в 2016 и 2018 годах, в сторону более низких – в 2017 году. Реакция растений на температурный фактор проявлялась в изменении сроков начала фенофаз: в 2017 году вегетация начиналась в более поздние сроки у 27 таксонов, сроки цветения задерживались у 43 таксонов на период от 7 до 30 дней, сроки плодоношения – у 38 таксонов на такой же период. Для 8 видов в 2017 году установлен более продолжительный период цветения.
В разные годы наблюдений для 12 видов выявлены близкие значения суммы среднесуточных температур на период начала бутонизации, для 10 видов – начала цветения, для 1 вида – начала плодоношения, что может свидетельствовать о тесной связи развития генеративной сферы этих таксонов с теплообеспеченостью.
Ежегодно образуют семена 35 таксонов, из них 14 дают самосев. Семенное размножение отсутствует или семена созревают только в отдельные годы у 7 таксонов.
Все исследованные виды предварительно оцениваются как перспективные хорошо адаптированные для выращивания в культуре в условиях южной Карелии. Большинство характеризуется целым рядом декоративных качеств, длительным периодом вегетации, продолжительным цветением. Особых рекомендаций для северных садов заслуживает группа стабильно декоративных весеннее-летне-зимнезеленых растений, включающая 16 таксонов.
Благодарности
Работа выполнена при поддержке гранта РФФИ 18-44-100002 р_а.
Литература
Борисова И. В. Ритмы сезонного развития степных растений и зональных типов степной растительности Центрального Казахстана // Труды БИН им. В. Л. Комарова. Сер. 3. Геоботаника. 1965. Вып. 17. С. 64—99.
Булыгин Н. Е., Довгулевич З. Н. Некоторые результаты математического анализа вековых фенологических рядов // Межвузовский сборник законченных научно-исследовательских работ. Вып. 2. Л., 1974. С. 36—40.
Карписонова Р. А. Травянистые растения широколиственных лесов СССР: экологофлористическая и интродукционная характеристика. М.: Наука, 1985. 205 с.
Кищенко И. Т., Потапова М. Н. Сезонный рост побегов представителей рода Acer (Aceraceae) в условиях интродукции // Ученые записки ПетрГУ. 2008. № 2 (92). С. 52—57.
Кищенко И. Т. Рост и развитие аборигенных и интродуцированных видов семейства Pinaceae Lindl. в условиях Карелии. Петрозаводск: Изд-во ПетГУ, 2000. 211 с.
Кищенко И. Т. Рост и развитие интродуцированых видов рода Tilia L. (Tiliaceae) в условиях Карелии // Вестник Северного (Арктического) федерального университета. Архангельск. 2015. № 4. С. 56—65.
Лантратова А. С. Сезонные изменения в цикле развития хвойных, отличающихся происхождением и степенью устойчивости к воздействию низких температур // Влияние термического фактора на сезонную ритмику растений. Материалы Всесоюзной конференции. М.: Изд-во АН СССР, 1979. С. 68—72.
Лантратова А. С., Барская Т. А., Сергеева М. Н. Морозоустойчивость пихт, интродуцированных в южной Карелии // Научные доклады высшей школы. Биологические науки. 1969. № 11. С. 88—94.
Лантратова А. С., Задорожная Г. Л. Интродукция двух видов дуба в южной Карелии // Бюлл. ГБС АН СССР. 1971. № 79. С. 33-37.
Лантратова А. С. Ритм роста и развития сеянцев лиственницы сибирской // Ученые записки Петрозаводского государственного университета. Петрозаводск: Издательство Петрозаводского государственного университета, 1962. Т. 10. № 1.
Летопись природы России: фенология. Материалы I Международной фенологической школы-семинара в Центрально-Лесном государственном природном биосферном заповеднике, 13-17 августа 2018 г. Великие Луки, 2018. 224 с.
Методика фенологических наблюдений в ботанических садах СССР. М, 1975. 27 с.
Минин А. А., Ранькова Э. Я., Буйволов Ю. А., Сапельникова И. И., Филатова Т. Д. Фенологические тренды в природе центральной части Русской равнины в условиях современного потепления // Жизнь Земли. 2018. Т. 40. № 2. С. 162—174.
Седельникова Л. Л. Виды рода Hosta (Hostaceae) при интродукции в Западной Сибири // Вестник КрасГАУ. 2012. № 11. С. 73—78.
Современное состояние фенологии и перспективы ее развития: Материалы Международной научно-практической конференции, посвященной 115-летию со дня рождения выдающегося советского фенолога В. А. Батманова, 17-18 декабря 2015 г., Екатеринбург. Екатеринбург: ФГБОУ ВПО «УрГПУ», 2015. 276 с.
Фирсов Г. А. Древесные растения ботанического сада Петра Великого (XVIII-XXI вв.) и климат Санкт-Петербурга // Ботаника: история, теория, практика (к 300-летию основания Ботанического института им. В. Л. Комарова Российской академии наук): Труды международной научной конференции. СПб.: Изд-во СПбГЭТУ «ЛЭТИ», 2014. С. 208—215.
Фомина Т. И. Биологические особенности зимнезелёных поликарпиков в лесостепной зоне Западной Сибири // Вестник Томского государственного университета. Биология. 2012. № 1 (17). С. 43—51.
Штанько А. В. К вопросу изучения фенологии рапса в условиях Карелии // Сезонное развитие природы. М.: АН СССР, Моск. филиал Геогр. об-ва СССР, 1975. С. 41—43.
Штанько А. В., Поташева Е. А. Феноспектр цветения некоторых медоносных растений в условиях Карелии // Пути адаптации растений при интродукции на Севере: Межвуз. сб. Петрозаводск: ПГУ, 1982. C. 74—79.
Штанько А. В., Штанько С. А. Лекарственные растения: учебное пособие. Петрозаводск, 1993. 253 с.
Bjorkman A. D., Elmendorf S. C., Beamish A. L., Vellend M., Henry G. H. Contrasting effects of warming and increased snowfall on Arctic tundra plant phenology over the past two decades // Glob Chang Biol. 2015. Vol. 21 (12). P. 4651—4661.
CaraDonna P. J., Iler A. M., Inouye D. W. Shifts in flowering phenology reshape a subalpine plant community // Proceedings of the National Academy of Sciences. 2014. Vol. 111 (13). P. 4916—4921.
Cleland E. E., Chuine I., Menzel A., Mooney H. A., Schwartz M. D. Shifting plant phenology in response to global change // Trends Ecol Evol. 2007. Vol. 22(7). P. 357—365.
Schulman L., Lehvävirta S. Botanic gardens in the age of climate change // Biodiversity and Conservation. 2011. Vol. 20. Is. 2. P. 217—220.
Stucky B. J., Guralnick R., Deck J., Denny E. G., Bolmgren K., Walls R. The Plant Phenology Ontology: A New Informatics Resource for Large-Scale Integration of Plant Phenology Data // Front Plant Sci. 2018. Vol. 9. Article 517. P. 1—12.
Wolkovich E. M., Cleland E. E. The phenology of plant invasions: a community ecology perspective // Frontiers in Ecology and the Environment. 2011. Vol. 9. Is. 5. P. 287—294.
