

Введение
Дождь и почвенная влага – независимые от растений источники воды, однако если их недостаточно, то всегда доступен воздух, из которого растение может получить росу, снизив температуру листьев, а не дожидаясь ненадежного тумана.
В предыдущих работах были приведены доказательства охлаждения поверхности растений ниже точки росы в широком диапазоне климатических условий (Прохоров, 2015а), для многих видов растений (Прохоров, 2015б; Карпун и др., 2015), что подтвердило выдвинутую ранее гипотезу (Прохоров, 2013), состоящую в том, что растения активно конденсируют атмосферную влагу на своей поверхности, за счет снижения температуры поверхности (TL) листьев ниже точки росы (TD), при температуре воздуха TA>TD, т. е. при отсутствии тумана. Эволюционное значение этого явления проявляется в морфологических, анатомических и физиологических адаптациях растений, способствующих увеличению эффективности конденсации и поглощения влаги.
Явление снижения температуры живых листьев растений описано более полувека назад (Lange & Lange, 1963), и обычно объясняется транспирационным охлаждением (Gates, 1968), физическая природа которого соответствует испарительному охлаждению. Однако существует значительная группа растений, использующих CAM-фотосинтез или «кислотный метаболизм толстянковых». Преимущество механизма CAM-фотосинтеза состоит в том, что растение может держать свои устьица закрытыми в жаркие дневные часы, существенно снижая потери воды через транспирацию, и запасать углекислый газ ночью, при значительно более выгодной температуре и влажности. Высокая эффективность использования воды САМ-растениями, по сравнению с фотосинтезирующими растениями С3 и С4, делает некоторые растения с высокой продуктивностью привлекательными альтернативами продовольственным и биоэнергетическим культурам на полузасушливых землях (Cushman, 2017). Можно предположить, что CAM-растения позволят оценить эффект совокупного транспирационного и радиационного охлаждения растения в ночное время.
Среди них есть представители семейства Asparagaceae, для которых атмосферная влага является важным источником воды, и часто имеющие розеточную форму роста (Рис. 1). Показана эффективность такой морфологии в отношении сорбции тумана (Martorell & Ezcurra, 2007). Общее количество тумана, перехваченного розеточными растениями, росло с увеличением общей площади листьев, за счет того, что многочисленные узкие листья максимизировали эффективность перехвата на единицу площади. Наблюдалась тенденция к развитию «синдрома узколистности», по мере приближения ареала вида к районам, где часто бывает туман.

Рис. 1. A. Драконовы деревья (Dracaena draco L., Asparagaceae) – редкие и лекарственные эндемики Канарских островов в ботаническом саду 'Viera-i-Klaviho', о-в Гран Канария. B. Сеянцы Dracaena draco в возрасте 7 лет в климатической камере. (фото А. Прохорова).
Эксперименты по поливу пазух листьев драконова дерева – Dracaena draco L. (Nadezhdina & Nadezhdin, 2017) подтвердили гипотезу о том, что драцены способны направлять атмосферную воду через пазухи своих листьев в стволовые ткани. Этот механизм представляет собой альтернативный способ поглощения воды растениями и, по мнению авторов, особенно важен в туманных районах засушливого и полузасушливого климата.
Также показано (Jura‑Morawiec, Marcinkiewicz, 2020), что поверхность листьев Dracaena draco может впитывать воду. Толстая базальная часть листа работает как резервуар для воды, объем которого изменяется при увлажнении растений. Зрелые листья розетки более смачиваемы, чем молодые. Вода может поглощаться как через адаксиальную, так и через абаксиальную поверхности.
Данные представления о "смачиваемости" поверхности листьев вида, адаптированного к аридным условиям, противоречат наблюдаемым явлениям формирования капель воды и стекания влаги на гидрофобных поверхностях растений, обусловленных "эффектом лотоса" (Barthlott & Ehler, 1977). Скорее, вертикальная розеточная форма и гидрофобная поверхность, позволяют влаге быстро стекать к пазухам, где и происходит поглощение воды (Nadezhdina & Nadezhdin, 2017).
Сравнение микроморфологии устьиц (Klimko, Wiland-Szymańska, 2008) группы видов драконовых деревьев (D. draco, D. cinnabarii, D. ombet и D. schizantha) показало различия в строении кутикулы и восковых структур. Из приведенных авторами фотографий видно, что замыкающие клетки у D. draco менее защищены. Следовательно, замыкающие клетки устьиц, как обладающие наиболее гидрофильной поверхностью, могут являться центрами образования микрокапель росы. Кроме того, устьица D. draco равномерно распределены рядами на адаксиальной и абаксиальной сторонах листа, что имеет существенное значение для конденсации и поглощения воды.
Осталось понять, что является главным источником влаги для драцены - ненадежный туман или роса, еженощно выпадающая на охлажденных листьях?
Объекты и методы исследований
Жарким летом 2018 года (Рис. 2) в Карелии сложились необходимые (сходные с весной на Канарах) условия для изучения циркадных ритмов температуры поверхности листьев драконова дерева – Dracaena draco L. – солнце, жара и наличие объекта исследований. В этих условиях (Рис. 3) туман наблюдался не каждую ночь, и был обычно кратковременным, а температура листьев снижалась ниже точки росы постоянно и особенно сильно после рассеивания тумана.
Таблица 1. Символы и термины, используемые в диаграммах и тексте
| Параметр | Легенды диаграмм | Символы в тексте и формулах |
| температура воздуха | Air Temperature, °C | TA |
| температура почвы | Soil temperature, °C | TS |
| точка росы | Dew point, °C | TD |
| температура стебля | Stem IR-temperature, °C | TSt |
| температура листьев | Leaf IR-temperature, °C | TL |
| относительная влажность воздуха | Relative Humidity, % | RH |
| отклонение температуры поверхности листа от точки росы | Leaf °C Dewpt. dist. | ΔTL-D; ΔTD-L |
| отклонение температуры поверхности стебля от точки росы | Stem °C Dewpt. dist. | ΔTSt-D |
| отклонение температуры воздуха от точки росы | Air °C Dewpt. dist. | ΔTA-D |
| отклонение температуры воздуха от температуры поверхности листа | Air - Leaf °C temp. dist. | ΔTA-L |
| уровень солнечной радиации | Solar Radiation, Вт/м2 |
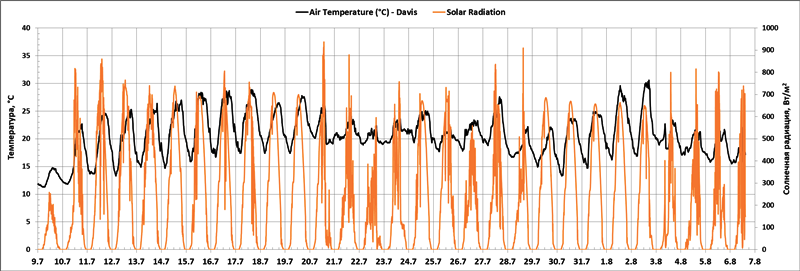
Рис. 2. Температура воздуха (°С) и уровень солнечной радиации (Вт/м2) по данным метеостанции Vantage Pro2 Plus в период с 9 июля по 7 августа 2018 г.
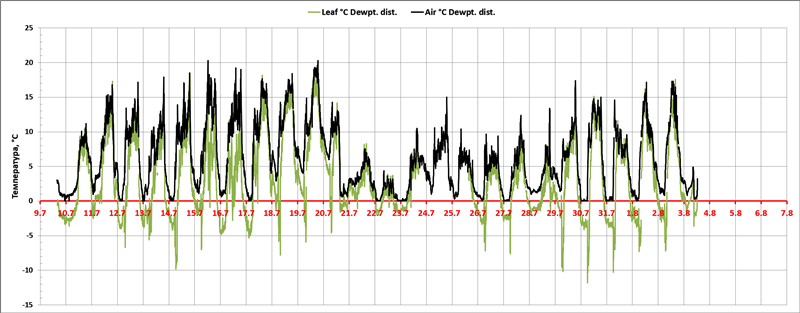
Рис. 3. Отклонение температуры воздуха и температуры поверхности листьев D. draco (ΔTA-D, ΔTL-D) от точки росы в период с 9 июля по 3 августа 2018 г.
Для исследований были выбраны два сеянца D. draco, высотой 60 и 80 см, культивируемые в контейнерах в климатической камере в Ботаническом саду Петрозаводского государственного университета с 2012 года (Рис.1B). Семена были собраны в районе Playa del Ingles (Gran Canaria, Spain) в 2011 году.
Полевые эксперименты
Исследования проводились с 9:00 24.08.2018 до 9:30 05.09.2018 под навесом из полиэтиленовой пленки для защиты от дождя. Полиэтилен был выбран в качестве защитного покрытия в связи с его прозрачностью в области окна прозрачности атмосферы 8 - 15 мкм, т.е. в диапазоне теплового излучения Земли.
Одно растение использовалось в период с 9 июля по 23 июля, второе с 25 июля по 3 августа 2018 г. За неделю до начала опыта и во время измерений полив растений не осуществлялся.
С помощью метеостанции Vantage Pro2 Plus (DAVIS Instruments), установленной непосредственно рядом с объектом исследований на высоте 3,5 метра над уровнем почвы, фиксировались следующие климатические параметры: температура воздуха - TA (°C); относительная влажность воздуха - RH (%); точка росы - TD (°C); количество осадков (мм/час); уровень солнечной радиации (Вт/м2) с интервалом времени 5 минут.
Данные о температуре (°C) воздуха - TA, почвы в контейнере - TS, поверхности листьев - TL, точке росы - TD, отклонения температуры поверхности листа от точки росы (ΔTL-D=TL-TD), а также, относительной влажности (%) воздуха - RH, получены с помощью инфракрасного термометра с интегрированным модулем влажности Testo 835-H1 (Testo) c выводом данных на компьютер с интервалом 5 минут. Параллельные измерения температуры поверхности стебля - TSt осуществлялись с помощью инфракрасного термометра DT-8855. В тестовых экспериментах отклонения показаний инфракрасных термометров друг от друга не превышали ±0,5°C.
ΔTL-D и остальные, приведенные в диаграммах, отклонения температур рассчитывались из экспериментальных данных.
Из результатов измерений исключены единичные ошибки в измерении RH и параметров, рассчитанных на ее основе (TD, ΔTL-D), вызванные порывами ветра или другими случайными факторами, действовавшими на протяжении одного 5-минутного цикла измерений и вызывавшие отклонения значений RH более чем на 10%.
При измерении TL и TSt использовался коэффициент эмиссии инфракрасных термометров - 0,95. Приборы располагались на высоте 0,6 метра от поверхности земли и были защищены от солнечного излучения пенопластовыми панелями. Расстояние до объекта 1 метр, диаметр пятна измерений – 20 мм. Выбор пятна измерений осуществлялся с помощью лазерных целеуказателей на обращенной к северу стороне растения.
В тексте, при описании наблюдаемых явлений, используются общепринятые названия или символы; в легендах на диаграммах используются обозначения, принятые в программном обеспечении "testo EasyClimate" для инфракрасного термометра Testo 835H1, или их аналоги для расчетных параметров или данных других приборов (таблица 1).
Эксперименты в климатической камере
Климатическая камера с полезным объемом 3,8 куб. метра и площадью 2 кв. метра, толщина стен и пола 10 см (пенопласт, стены покрыты изнутри влагостойкой зеркальной пленкой, пол – пленкой серого цвета), накрытая сверху стеклопакетом толщиной 2 см. Над стеклопакетом установлено 4 пары светодиодных светильников УСС БИО с максимумами излучения на 660 и 450 нм, включение и отключение которых осуществлялось последовательно, согласно установленной программе с интервалом 1,5 часа. Продолжительность светового дня – 12 часов. TA и RH менялись спонтанно, сопряженно с изменением освещенности. Во время эксперимента TA в помещении, до включения освещения климатической камеры, составляла 9-10°C. Освещенность контролировалась в верхней точке кроны растения с помощью люксметра ET-965 (Flus). Измерения осуществлялись с помощью инфракрасного термометра с интегрированным модулем влажности Testo 835-H1 (Testo) и инфракрасного термометра DT-8855 для параллельных измерений TSt.
Загрузка данных с приборов производилась программными средствами производителей оборудования. Построение диаграмм осуществлялось с помощью MS Excel.
Результаты и обсуждение
Открытый грунт
Для анализа полученных результатов был выбран период с 29 июля по 3 августа (Рис. 3) – пять идеальных солнечных дней и звездных ночей. Восход солнца 29.07.2018 – 4:35, заход – 21:25, по данным датчиков солнечной радиации.
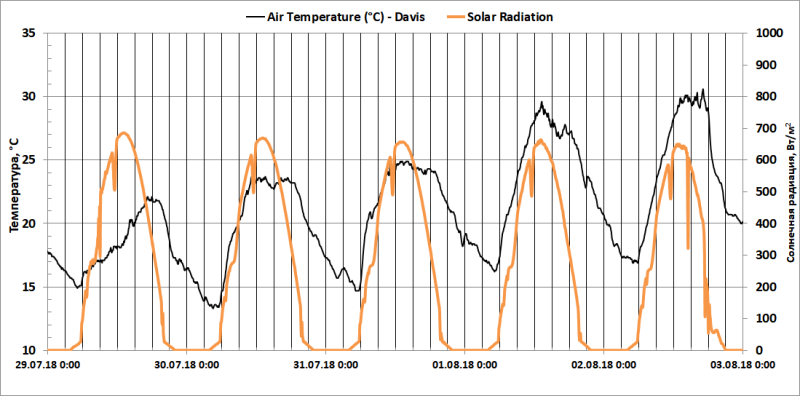
Рис. 4. Температура воздуха (°С) и уровень солнечной радиации (Вт/м2) по данным метеостанции Davis Vantage Pro2 Plus в период с 29 июля по 3 августа 2018 г.
Данные метеостанции (Рис. 4) демонстрируют постепенное увеличение TA по мере прогрева нижних слоев атмосферы. Однако непосредственно рядом с растениями (Рис. 5) наблюдаются максимумы TA в 7 и 19 часов и TS в 9 часов утра. Боковое солнечное освещение на рассвете и на закате вызывало быстрый нагрев припочвенного слоя воздуха и контейнеров с растениями.
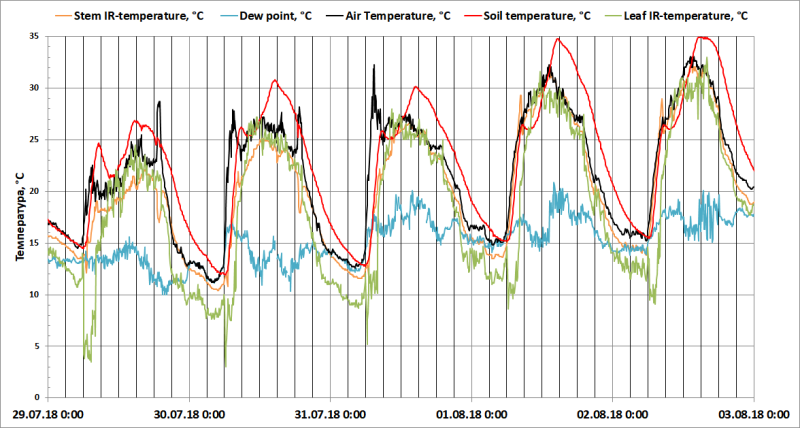
Рис. 5. Точка росы (°С), температуры воздуха на высоте кроны объекта, почвы в контейнере, листьев (по данным инфракрасного термометра Testo 835-H1) и стебля Dracaena draco (по данным инфракрасного термометра DT-8855).
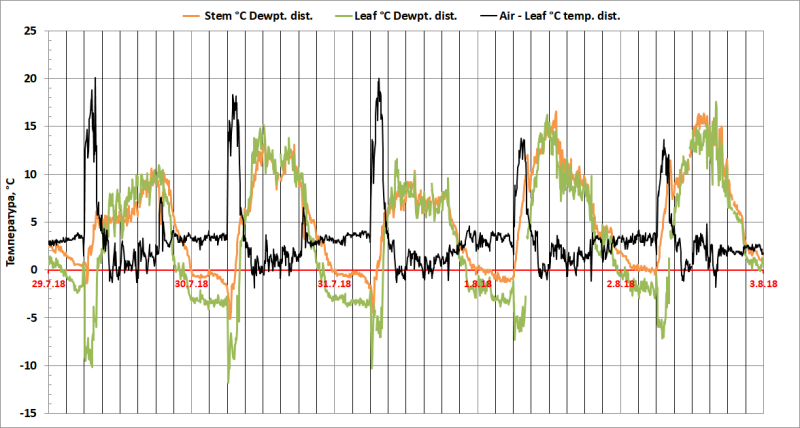
Рис. 6. Отклонения температуры поверхности листьев и стебля Dracaena draco от точки росы (∆TL-D = TL–TD, ∆TSt-D = TSt–TD) и отклонения температуры воздуха от температуры листьев (∆TA-L = TA–TL).
∆TA-L составляла около 0-1°С с 10:00 до 18:00, 3-4°С с 22:00 до 6:00, 13-20°С с 6 до 9 часов утра (Рис. 6).
Основной параметр, интересующий нас в данном исследовании – ∆TL-D, разница между температурой поверхности листа (TL) и точкой росы (TD). Именно он определяет количество росы, которая выпадает на поверхности листа (Рис. 6) в единицу времени. И этот параметр ведет себя с одной стороны предсказанно, а с другой – удивительно.
Ожидаемо, что TD>TL ночью, когда устьица у CAM-растения открыты и на тепловое излучение накладывается испарительное охлаждение. Однако в 6 утра, при быстром увеличении TA, вызванной ростом солнечной радиации, TL стремительно снижается еще на 10-17°С, снижая TL на 7-8°С ниже TD с 6 до 9 часов утра. На 4 день эффект уменьшается, но не исчезает.
Совпадение максимума TA и минимума TL, позволяет считать испарение росы с поверхности листьев, наиболее вероятной причиной сильного охлаждения листьев в утренние часы. Совпадение указанных экстремумов приводит к значительному увеличению выпадения росы в это время, компенсирующему затраты воды на испарение.
Результаты опытов в открытом грунте не позволяют сделать однозначный вывод о соотношении физических механизмов охлаждения листьев D. draco, для этого необходимо сопоставить их с результатами исследований в климатической камере.
Климатическая камера
На диаграмме (Рис. 7) приведены циркадные ритмы TD (°С), TA, TS, TL и TSt Dracaena draco в период с 7.03 по 13.03 2018.
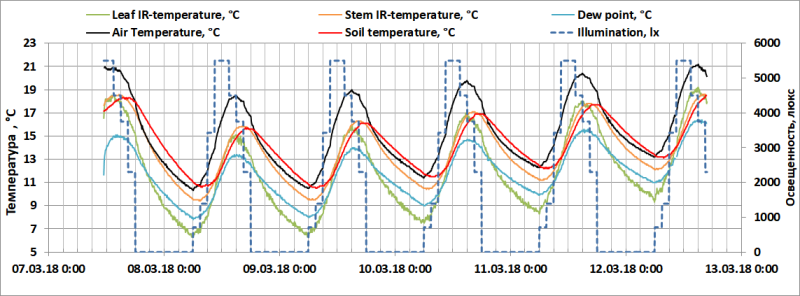
Рис. 7. Освещенность (люкс), точка росы (°С), температура воздуха, почвы в контейнере, листьев (по данным инфракрасного термометра Testo 835-H1) и стебля Dracaena draco (по данным инфракрасного термометра DT-8855).
Ключевым отличием климатической камеры от открытого грунта является практически полное отсутствие естественного теплового излучения, что не позволяет растениям остывать столь же интенсивно, как в полевом эксперименте, когда ∆TL-D= -3°С. В ночное время температура поверхности стеклопакета снижается до температуры воздуха в помещении (~10°С) и в климатической камере, где TA снижается медленнее, создаются условия для незначительного теплового излучения с поверхности растений.
Даже в этих условиях, в ночное время, создается длительный (~15 часов) период самоорошения D. draco (Рис. 8), когда ∆TL-D достигал 0°С через 1,5 часа после выключения света при TL~12-14°С и снижался до ∆TL-D= -1,5°С.
Кроме того, в климатической камере светодиодное освещение не дает всплеска инфракрасного излучения, сопутствующего началу светового дня в открытом грунте. В данных условиях экстремальное снижение TL, наблюдавшееся в открытом грунте с 6:00 до 9:00, после включения освещения в 6:00 в климатической камере не обнаружено. Характер изменений температурных характеристик среды (TA, TS) был монотонным, и следовал за искусственным суточным ритмом освещенности и температуры. Таким образом, получено косвенное подтверждение интенсивного испарительного охлаждения растений в начале светового дня на открытом воздухе при высоком уровне выпадения росы в ночное время.
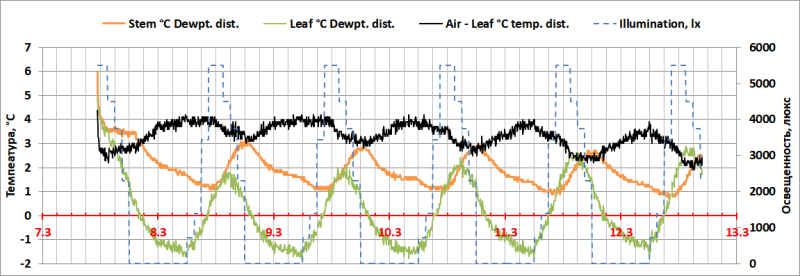
Рис. 8. Отклонения температуры поверхности листьев и стебля Dracaena draco от точки росы (∆TL-D=TL–TD) и отклонения температуры воздуха от температуры листьев (∆TA-L=TA– TL) в климатической камере.
Расчет количества росы
В 2019 году нами опубликованы результаты экспериментальных исследований (Прохоров, Пяскин, 2019) в результате которых получены значения коэффициента конденсации (KC) воды на охлажденной поверхности термоэлемента, позволяющие количественно оценить эффективность самоорошения растений. Для оценки количества конденсируемой влаги на поверхности растений необходимы данные о площади поверхности (SL), отклонениях температуры поверхности от точки росы (∆TD-L=TD–TL) и продолжительность периода времени при которой фиксируются отрицательные значения данного параметра.
Расчет объема выпадающей росы (VD) велся на основе определенных нами коэффициентов конденсации (Прохоров, Пяскин, 2019) по формуле:
VD = KC × ∆TD-L × t × SL
— где коэффициент конденсации, KC = 1,25 мкл/(см2 × час × град) – для условий RH≥80% и TA ≤20°C; ∆TD-L - отклонение температуры поверхности от точки росы; t - продолжительность времени конденсации (период измерения 5 минут); SL – площадь поверхности листьев.
Согласно (Marrero, Almeida Perez, 2012) длина листа D. draco составляет 79,7±13,02 см, а ширина 3,3±0,4 см, что дает среднюю суммарную площадь абаксиальной и адаксиальной поверхности одного листа (SL) ~500 см2. На диаграмме (Рис. 9) приведены результаты расчета количества росы, выпадающей на 1 м2 поверхности (~20 листьев D. draco) каждые 5 минут в открытом грунте.
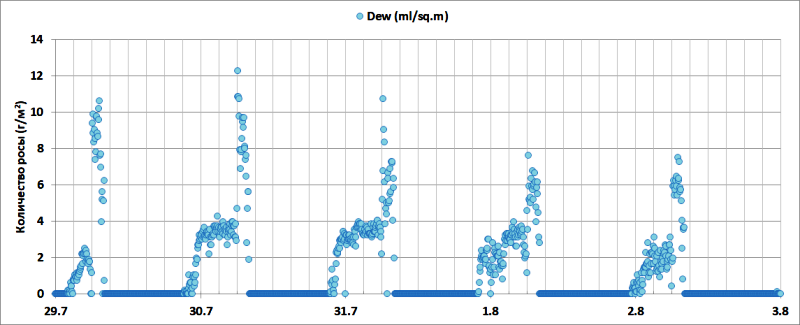
Рис. 9. Расчетное количество росы, выпадающей на 1 м2 поверхности листьев в течение 5 минут.
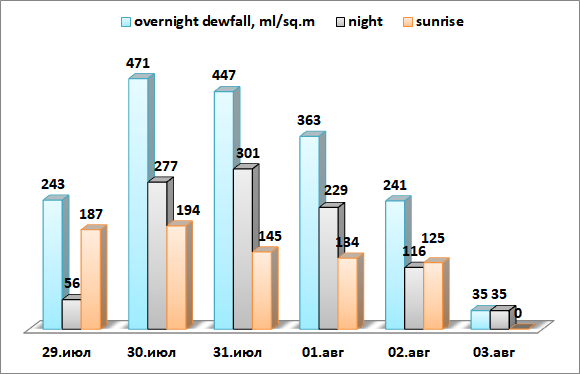
Рис. 10. Расчетное количество росы (мл), выпадающей за сутки на 1 м2 поверхности листьев, в т.ч. в ночное время и в утренние часы.
Общий объем росы (рис.10) состоит из двух компонент: при ясной погоде с 22:00 до 6:00 утра в интервале ΔTD-L 0-4°C конденсируется более половины всего объема росы, с 6:00 до 9:00 в интервале ΔTD-L 0-10°C выпадает от 1/2 до 2/3 ночного объема.
В климатической камере при ΔTD-L=1,5°C растение конденсирует около 200 мл росы за 12-14 часов, с максимальным выпадением росы 1,8 мл/м2 за 5 минут (рис.11).
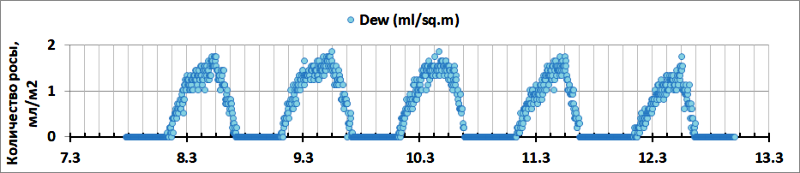
Рис. 11. Расчетное количество росы, выпадающей на 1 м2 поверхности листьев в течение 5 минут в климатической камере.
Выводы и заключение
- На примере драконова дерева (Dracaena draco L.) показано, что CAM-растение в аридных условиях способно конденсировать и поглощать в течение ночи воду, добываемую за счет снижения температуры листьев на 3-4°С ниже точки росы с 22:00 до 6:00.
- Сопоставление результатов исследования в открытом грунте и в климатической камере не позволяют сделать однозначный вывод о вкладе теплового излучения и транспирационного охлаждения в снижение температуры листьев. Можно предположить, что в обоих случаях в ночной период имеет место транспирация САМ-растения, охлаждающий эффект которой в открытом грунте усилен тепловым излучением.
- В ходе экспериментов получены данные, свидетельствующие о существовании особого периода в начале светового дня, когда температура поверхности листьев снижается на 5-10°C под открытым небом, сопряженно с быстрым ростом температуры воздуха. В климатической камере такой период отсутствует. Совпадение максимума TA и минимума TL позволяет считать испарение росы с поверхности листьев наиболее вероятной причиной сильного охлаждения листьев в утренние часы. Совпадение указанных экстремумов приводит к значительному увеличению выпадения росы (в 3 раза), компенсирующему затраты воды на испарение.
- Рассчитанное количество росы конденсируемое каждым квадратным метром поверхности листьев драконова дерева после захода солнца, составляет около 0,3 - 0,5 литра за 11 часов при TA в диапазоне 12-15°C и RH>80% в открытом грунте.
Автор всегда недоумевает, когда устьица рассматриваются как орган, отвечающий исключительно за газообмен. В тех случаях, когда устьица открыты, а поверхность листа покрыта росой (дистиллированной водой), они будут функционировать как орган, поглощающий воду по градиенту осмотического давления. Как видно из приведенных экспериментальных данных (Рис. 6, Рис. 8) для САМ-растений такая ситуация типична в ночное время.
А если учесть, что замыкающие клетки у D. draco менее защищены восками и расположены рядами (Klimko, Wiland-Szymańska, 2008), то они, как обладающие наиболее гидрофильной поверхностью, могут являться центрами образования микрокапель росы. Вода поглощается по градиенту осмотического давления устьицами и вполне вероятно, что ночью невозможно наблюдать формирование крупных капель воды. Потерями воды на испарение в ночное время можно пренебречь, следовательно, роса, выпадающая в ночной период, поглощается листьями драконова дерева.
Роса, выпадающая в утренние часы при быстром росте температуры, в значительной степени испаряется, однако ее количество сильно возрастает за счет роста градиента температуры между поверхностью листа и точкой росы. В это время устьица САМ-растения должны быть закрыты. Формирующиеся на поверхности замыкающих клеток капли росы быстро увеличиваются в размере и роса стекает в пазухи листьев, как и капли росы, формирующиеся на участках поверхности, лишенных устьиц.
Использование всей охлажденной поверхности листа для конденсации атмосферной влаги и поглощение выпавшей росы с помощью открытых в ночное время устьиц и поглощающих воду тканей в пазухах листьев - поразительная адаптация к условиям недостатка водных ресурсов, включающая морфологические, анатомические и физиологические приспособления, позволяющие целесообразно использовать многочисленные законы природы и уникальные свойства воды.
Явление самоорошения растений может быть использовано для повышения урожайности культивируемых CAM-растений – ананас, агава, алоэ и др. рост которых связан с накоплением воды в тканях. Оседающая ночью и утром на листьях холодная роса может являться хорошим источником СО2 для растений, ввиду его большей растворимости при низких температурах.
Благодарности
Это исследование не получило специального гранта от какого-либо фонда в государственном, коммерческом или некоммерческом секторах. Вместе с тем автор искренне благодарен Министерству науки и образования РФ за субсидии 2016-2018 гг., которые позволили нам закупить оборудование, необходимое для научных исследований.
Литература
Карпун Ю. Н., Коннов Н. А., Кувайцев М. В., Прохоров А. А. Активная конденсация атмосферной влаги как механизм самоорошения почвопокровных растений // Hortus bot. 2015. Т. 10, с. 11–17. URL: http://hb.karelia.ru/journal/article.php?id=2802. DOI: 10.15393/j4.art.2015.2802
Прохоров А. А. Точка росы - неизученный фактор в экологии, физиологии и интродукции растений // Hortus bot. 2015b. Т. 10,., С.4–10. http://hb.karelia.ru/journal/article.php?id=2801. DOI: 10.15393/j4.art.2015.2801
Прохоров А. А. Оптимальные климатические условия для конденсации атмосферной влаги на поверхности растений // Hortus bot. 2015a. Т. 10, с. 18–24. URL: http://hb.karelia.ru/journal/article.php?id=3143 , https://doi.org/10.15393/j4.art.2015.3143
Прохоров А..А. Активная конденсация воды растениями // Принципы экологии. ПетрГУ. 2013. № 3. С. 58—61. DOI: 10.15393/j1.art.2013.2921.
Barthlott, W., & Ehler N. (1977). Raster-Elektronenmikroskopie der Epidermis-Oberflächen von Spermatophyten. Tropische Und Subtropische Pflanzenwelt, p. 105.
Cushman J. (2017) CAM Plants in Encyclopedia of Applied Plant Sciences (Second Edition) Vol. 1, pp. 60-77.
Gates D. M. (1968). Transpiration and Leaf Temperature. Annual Review of Plant Physiology, 19, 211–238.
Jura Morawiec J., Marcinkiewicz J. (2020) Wettability, water absorption and water storage in rosette leaves of the dragon tree (Dracaena draco L.) // Planta 252:30 https://doi.org/10.1007/s00425-020-03433-y
Klimko M., Wiland-Szymańska J. (2008): Scanning electron microscopic studies of leaf surface in taxa of genus Dracaena L. (Dracaenaceae). Rocz. AR Pozn. 387, Bot.-Stec. 12: 117-127.
Lange O. L., & Lange R. (1963). Untersuchungen uber Blattemperaturen, Transpiration und Hitzeresistenz an Pflanzen mediterraner Standorte (Costabrava, Spanien). Flora, 153, 387–425.
Marrero A., Almeida Perez R.S. (2012) A new subspecies, Dracaena draco (L.) L. subsp. caboverdeana Marrero Rodr. & R. Almeida (Dracaenaceae) from Cape Verde Islands // International Journal of Geobotanical Research, Vol. N.2. pp. 35-40
Martorell C., & Ezcurra E. (2007). The narrow-leaf syndrome: A functional and evolutionary approach to the form of fog-harvesting rosette plants. Oecologia, 151(4), 561–573. https://doi.org/10.1007/s00442-006-0614-x
Nadezhdina N., & Nadezhdin V. (2017). Are Dracaena nebulophytes able to drink atmospheric water? Environmental and Experimental Botany, 139, 57–66. https://doi.org/10.1016/j.envexpbot.2017.04.005
