

Введение
Виды рода Larix Mill. относятся к деревьям выдающегося интереса для лесоводов (Уханов, 1949). Большинство из 10-15 известных видов этого рода введены в культуру и используются в странах холодного и умеренного климата в коммерческих целях, в лесном, лесопарковом хозяйстве и озеленении. Некоторые виды доказали свою пригодность для разведения в более тёплых странах. Однако, поскольку это преимущественно виды высоких широт и холодных областей земного шара, их культура в условиях более мягкого климата делает их уязвимыми к весенним заморозкам, а также появляются проблемы с различными болезнями и вредителями (Dallimore et al., 1966; Grimshaw, Bayton, 2009). В Ботаническом саду Петра Великого в Санкт-Петербурге лиственницы были одними из первых интродуцентов. Здесь они зарекомендовали себя как самые зимостойкие древесные растения.
В начале XX в. в парке-дендрарии Ботанического сада Петра Великого Ботанического института им. В. Л. Комарова РАН (БИН) в Санкт-Петербурге в последние годы заметно возросло усыхание и гибель древесных растений. Почвы Ботанического сада БИН были обследованы на присутствие оомицетов из рода Phytophtora в начале 1990-х гг., ни в одном почвенном образце, взятом из ризосферы древесных растений, фитофторы тогда не обнаружилось. В результате исследования ризосферной почвы больных и здоровых растений, проведенного в 2011-2015 гг. было обнаружено широкое распространение этих фитопатогенов. Это почвообитающие корнепоражающие виды, представляющие большую опасность для произрастающих в парке деревьев и кустарников. Фитофтора обнаружена на деревьях, кустарниках и лианах 31 вида, в том числе видах рода Larix. В образцах из ризосферы Larix decidua на участке 140 в 2012-2013 гг. обнаружен вид Phytophthora cactorum. Дерево полностью засохло и к настоящему времени удалено. Корневые патогены видов рода Phytophthora из группы грибоподобных оомицетов, являются инициирующими биотическими факторами, способными активно поражать древесные растения и кустарники, вызывая корневую гниль и усыхание растения. При этом Ph. citricola, Ph. plurivora и Ph. quercina были впервые отмечены в Российской Федерации. Вид Ph. quercina до этого был известен в Центральной и Южной Европе и на широте Санкт-Петербурга до этого не отмечался (Веденяпина и др., 2014а,б; Веденяпина, Фирсов, 2014; Фирсов и др., 2014; Веденяпина и др., 2015).
Паразитическая активность видов рода Phytophthora во многом определяется внешними факторами среды и усиливается на фоне потепления климата (Фирсов, 2014). Изменение климатических факторов, главным образом температуры и влажности, в сторону создания неблагоприятных для растений условий приводит к ухудшению состояния деревьев и активизации патогенов. Распространению фитофтор способствует возрастание количества осадков, ослабление морозов, увеличение вегетационного сезона и летних температур, сокращение зимнего периода и связанное с этим уменьшение промерзания почвы. По мнению Е. Г. Веденяпиной и др. (2014а), столь быстрое расширение ареалов почвообитающих фитофтор и их адаптационная способность в настоящее время можно определить двумя основными факторами. Это резкое увеличение международной торговли растениями, латентное поражение корневой системы которых фитофторами может быть незаметным. А также существенное и быстрое изменение климата с повышением температуры, увеличением осадков и повышением влажности воздуха и почвы неизбежно воздействуют на активность, жизненный цикл и сохранение патогенных организмов.
В разных странах мира фитофторы напоминают о себе ежегодно мощными и часто неожиданными вспышками болезней растений, носящими характер эпифитотий и панфитотий, уничтожая целиком не только урожаи сельскохозяйственных культур, но деревья и целые экосистемы в природе. Поскольку погибшие древесные растения не несут каких-либо признаков инфекции (то есть, спороношений, плодовых тел и пр.), их гибель часто объясняется абиотическими факторами или физиологическими причинами – вымоканием, засухой, обморожением, воздействием высоких и слишком низких температур, загрязнением воздуха, кислотными дождями, нехваткой микроэлементов, азота, фосфора и т.п. На самом деле это часто результат деятельности почвообитающих фитофтор.
Таким образом, если ранее в Санкт-Петербурге в XVIII-XX вв. основным фактором, ограничивающим культуру древесных растений, была недостаточная морозостойкость, то в последние всё более заметным и важным становится воздействие болезней и вредителей. При обычных визуальных фенологических наблюдениях причины корневых гнилей и гибели растений определить трудно или невозможно. Изреживание кроны, суховершинность, изъязвление ствола и ветвей, появление черных пятен, хлороз, внезапное усыхание, гниль корней и корневой шейки – симптомы, которые стали все чаще наблюдаться в Саду. Подобная патология древесных пород обычно объясняется абиотическими факторами. Однако такие же симптомы характерны и для болезней растений, вызванных корнепоражающими почвообитающими оомицетами из рода Phytophthora. Считается, что более 66% болезней тонких корней и более 90% всех гнилей корневой шейки вызываются видами рода Phytophthora (Jung, 2006). Настоящее исследование является продолжением начатого в 2011-2012 гг. изучения причин усыхания деревьев и кустарников на территории Ботанического сада Петра Великого Ботанического института им. В. Л. Комарова РАН (Веденяпина и др., 2014а,б).
Объекты и методы исследований
Обследование древостоя. Материалом для исследования служили растения лиственницы (Larix Mill.) Ботанического сада Петра Великого Ботанического института им. В. Л. Комарова РАН в Санкт-Петербурге. Один из авторов статьи, Г. А. Фирсов, является куратором дендропитомника с 1986 г. и парка-дендрария с 1994 г., с этих лет проводится непрерывный мониторинг за древесными растениями коллекции открытого грунта. Ежегодно отмечается общее состояние деревьев, повреждения, наличие дупел, трещин и морозобоин, соотношение засохших и живых ветвей в кроне, наклон ствола и особенности кроны, наличие плодовых тел грибов и гнилей при особом внимании к корневой шейке дерева. При определении категории состояния использовалась лесопатологическая методика (Мозолевская и др., 1984), с подразделением на 6 категорий состояния:
1 (без признаков ослабления); 2 (ослабленные); 3 (сильно ослабленные); 4 (усыхающие); 5 (сухостой текущего года); 6 (сухостой прошлых лет).
Естественная периодизация года (календарь природы) принята по Н. Е. Булыгину (1982). Фенологические наблюдения проводились по методике Н. Е. Булыгина (1979). Для оценки обмерзания использовалась шкала П. И. Лапина (1967): 1 – отсутствие повреждений… 7 – гибель растения с корнем. Для анализа тепло-влагообеспеченности использованы данные метеостанции Санкт-Петербургского Северо-Западного территориального управления по гидрометеорологии и контролю природной среды с региональными функциями. Биометрические измерения выполнены в 2015 г. Высота растений до 3,00 м измерялась мерной нивелирной рейкой с точностью до 1 см, до высоты 5,30 м – с точностью до 0,1 м. Высота более крупных деревьев – лазерным высотомером Nikon Forestry Pro с точностью до 0,2 м. Диаметр ствола (см) измерялся на высоте груди (1,3 м). Представляется возможным также сравнить данные по размерам и краткой характеристике для растений вида рода Larix, приведённые Северо-Западным лесоустроительным предприятием Всесоюзного объединения "Леспроект" в 1981 г. ("Инвентаризационное описание Ботанического института им. Комарова АН СССР, г. Ленинград") с данными, полученными при обследовании и обмерах каждого дерева современной коллекции. По инвентаризации Сада, проведенной в 1981 г. "Леспроектом", тогда выделялось три категории состояния: хорошее (хор.), удовлетворительное (уд.) и неудовлетворительное (неуд).
Отбор почвенных проб. В начале октября 2015 г., в конце первого феноэтапа подсезона "Золотая осень" производилось обследование участков с наиболее сильным усыханием на территории Сада. В том числе 3 образца были отобраны под деревьями лиственницы: Larix decidua (участок 140) и L. sibirica (участок 3, 128). Отбор почвенных проб осуществлялся с помощью стальных цилиндров диаметром 5 см и длиной 20 см. Точки отбора проб были приурочены к отдельным взрослым деревьям с расстоянием от 10 до 50 см от ствола в проекции кроны (в среднем пробы отбирались на расстоянии 15–30 см). Перед взятием пробы самый верхний неразложившийся слой подстилки удалялся. Дальнейший анализ собранных образцов включал методы детекции и идентификации патогена.
Изоляция фитофтор методом приманок. Постановка эксперимента проводилась в лабораторных условиях на базе БИН в лаборатории систематики и географии грибов. Часть почвы (20 г) из каждого образца помещали в отдельные пластиковые боксы и насыщали дистиллированной водой таким образом, чтобы над поверхностью почвы находилось не менее 2 см воды. Почву хорошо взбалтывали и давали отстояться до полной прозрачности поверхностного слоя воды. На поверхности воды раскладывали приманки - лепестки красной гвоздики. Выход патогена на приманку отслеживали путем визуального наблюдения и микроскопирования приманок. Обесцвечивание лепестков гвоздики указывало на возможность поражения живых тканей оомицетами. Обесцвечивание началось на 4-й день эксперимента (9 октября). Световое микроскопирование подтвердило наличие патогена в лепестках гвоздики. Были обнаружены многочисленные зооспорангии на приманках в пробах, в том числе в почвенной пробе, отобранной под Larix decidua. Участки лепестков, на которых достоверно имелся мицелий или зооспорангии, затем замачивались в лизис-буфере (2% СТАВ) на несколько дней.
Выделение ДНК, амплификация, секвенирование последовательностей и анализ молекулярных данных. Изоляция ДНК осуществлялась из материала с помощью NecleoSpin Plant II Kit (Macherey-Nagel) согласно прилагаемому протоколу. Для амплификации участка гена цитохромоксидазы COX2 (мДНК) использовались праймеры FMPhy-8b и FMPhy-10b (Martin et al., 2007). Секвенирование полученных участков производилось на автоматическом секвенаторе ABI 3130 (Applied Biosystems, USA) с использованием меченых реактивов BigDyeTM Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems) и той же пары праймеров. Главной целью проведения молекулярного анализа была видовая идентификация образца, а также сравнение полученной нуклеотидной последовательности COX2-региона с последовательностями близких видов, взятых из базы GenBank (National Center for Biotechnology Information, 2015).
Принятые далее сокращения: вег. – в вегетативном состоянии, выс. – высота, диам. – диаметр, окрест. – окрестности, пл. – плодоносит, пос. – посадка (год высадки на постоянное место из питомника в Парк), разн. – разновидность, уч. – участок, ф. – форма, шир. – ширина, экз. – экземпляр.
Результаты и обсуждение
В Ботаническом саду Петра Великого Ботанического института им. В. Л. Комарова РАН выращиваются следующие виды и формы лиственницы.
Larix archangelica Laws. - Лиственница архангельская. 9 экз., уч. 24, 29 (2 шт.), 30, 33 (2 шт.), 54, 59, 60. Пять деревьев на уч. 24 (№ 2), 29 (№ 2 и 14), 30 (№ 1), 33 (№ 22) представляют наиболее старые деревья в аллейных посадках 1820-х гг. (раньше не выделялась из L. sibirica). Уч. 60: семена из природы Архангельской обл., Пинежский район, окрест. посёлка Голубино, всх. 2005 г., пос. 2012 г. – самый быстрорастущий из молодых экз., 4,38 м выс. в 11 лет. Уч. 59: то же, пос. 2013 г. Уч. 54: семена из природы Архангельской обл., Плесецкий район, окрест. посёлка Озерское, всх. 2005 г., пос. 2013 г. Второе, молодое, дерево на уч. 33 пос. в 2015 г.: семена из Архангельской обл., Голубино, всх. 2005 г. Пл. Именно этот вид образует старейшие лесные культуры в Линдуловской роще на Карельском перешейке в Ленинградской обл. (с 1738 г.).
Larix cajanderi Mayr - Лиственница Каяндера. В Ботаническом саду БИН представлена молодыми растениями из природных местообитаний российского Дальнего Востока, привезенных сотрудниками Сада. 5 экз. Уч. 94 и 132 (2 экз.): растение из экспедиции Сада на Камчатку, окрест. Козыревска, у подножья вулкана Толбачик, 300 м н.у.м., в 2000 г., пос. 2009 г. (уч. 94) и 2010 (уч. 132). Уч. 60: растение из экспедиции Сада на остров Сахалин, гора Вайда, 400 м над уровнем моря, в 2004 г., пос. 2013 г. Уч. 33: пос. 2015 г.: семена из окрест. Магадана, всх. 2005 г. Пл. Может расти на холодных, сырых и бедных почвах, при очень низких зимних температурах воздуха.
Larix czekanowskii Szafer (L. sibirica × L. dahurica) - Лиственница Чекановского. 3 экз., уч. 14, 83, 117. Очевидно, в ботаническом саду БИН лиственница Чекановского впервые введена в культуру, хотя как самостоятельный таксон известна лишь с 1913 г. (Фирсов, Орлова, 2008). Деревья на уч. 14 ( 20) и 117 отмечены в путеводителе В.В. Уханова (1936). Возраст самого старого дерева на уч. 14 более 180 лет – то есть, всх. около 1830 г., оно же самое крупное – 26,8 м выс. и 84 см диам. Пл.
Larix dahurica Laws. (L. gmelinii (Rupr.) Rupr.) - Лиственница даурская. 14 экз., уч. 14, 27, 58, 82, 129, 130. Второй по численности вид в парке после лиственницы сибирской. Старые деревья достигают такого же возраста, около 200 лет, как и лиственницы сибирские. Экз. на уч. 26 и 27 еще в начале XX в. относились к замечательным деревьям парка (Липский, 1913). В Саду появилась до 1820 г. (Связева, 2005) и представлена здесь постоянно и без перерывов по настоящее время. Пл., образует самосев. Самый крупный экз. по высоте (уч. 14 № 20): 31,6 м, по диаметру ствола (уч. 27 № 5): 113 см. Введена в культуру Ботаническим садом БИН (Фирсов, Орлова, 2008). По мнению A. Rehder (1949), дата интродукции – 1827 г., однако в Санкт-Петербурге появилась раньше этой даты. На большей части своего обширного ареала связана с вечной мерзлотой и болотами.
Larix dahurica Laws. f. fastigiata Sr. - Лиственница даурская, ф. пирамидальная. 2 экз., уч. 55. Семена из природы от С. Н. Горошкевича: Баргузинский заповедник, у озера Байкал (отбор из сеянцев). Всх. 2006 г., пос. 2013 г. Лучший экз. в возрасте 10 лет 2,45 м выс. Вег.
Larix decidua Mill. - Лиственница европейская. 11 экз., уч. 14, 32, 48, 92, 94, 140, 145. Деревья почти все старые, до 200-летнего возраста. Самая первая лиственница, известная в Саду, выращивалась ещё в 1793 г., позже Ф. Б. Фишер (1837) испытал её повторно в 1833 г. и признал зимостойкой, с тех пор растёт в парке постоянно. Лучшие два дерева в аллее на уч. 48 – старые, толстые и высокие, пос. до 1835 г. (Связева, 2005). Уч. 140: засыхающий экз., ещё не достигший предельного возраста. Из молодых посадок – дерево на уч. 32 № 21: семена из природы Италии, всх. 1984 г., пос. 18.09.1992. Дерево на уч. 48 (№ 23) самое толстое среди лиственниц Парка, вместе с деревом лиственницы даурской на уч. 27 – 113 см. (превосходя толщину деревьев в Линдуловской роще). Самое высокое дерево этого вида (уч. 48 № 24): 30,0 м выс. Пл. В отличие от лиственницы даурской, плохо реагирует на заболачивание. Устойчива к городскому задымлению и отличается быстрым ростом. Для этого вида характерно более продолжительное охвоение на протяжении вегетационного сезона по сравнению с другими лиственницами, главным образом, из-за более позднего окончания вегетации.
Larix decidua Mill. subsp. polonica (Racib. ex Woycicky) Domin - Лиственница польская. 3 экз., уч. 123. В Саду первый раз испытана в 1956 г., однако вначале просуществовала лишь до 1967 г. (Связева, 2005). В современной коллекции один образец, группа из трёх деревьев: семена из Польши (Краков, институт фармакологии), всх. 1991 г., пос. в 2001 (2 шт.) и 2002 г. (1 шт.). Лучший экз. 16,5 м выс. в возрасте 25 лет. Пл. Как и лиственница европейская, поздно оканчивает вегетацию. В природе находится под угрозой исчезновения, была занесена в Красную книгу СССР (растёт в Карпатах и в районе реки Висла, в горах).
Larix decidua Mill. f. microcarpa Beissn. - Лиственница европейская, ф. мелкошишечная. Два невысоких дерева у забора вдоль набережной Большой Невки (уч. 35). Пл. В Саду появилась до 1950 г. (Связева, 2005). Очевидно, в культуре встречается очень редко.
Larix decidua Mill. f. pendula (Laws.) Henk. et Hochst. - Лиственница европейская, ф. плакучая. 1 экз., уч. 94. Даже для типичной лиственницы европейской свойственны тонкие и длинные повислые побеги. Однако встречаются деревья с особенно выраженной плакучестью кроны, которые относят к f. pendula. Известна с 1836 г. (Фирсов, Орлова, 2008). Иногда такие деревья можно встретить в садах и парках Санкт-Петербурга. По данным инвентаризации 1981 г. возраст дерева на уч. 94 сейчас около 85 лет. Пл.
Larix decidua Mill. f. pendulina Regel - Лиственница европейская, ф. ползучая. 5 экз., уч. 94. Группа из трёх искривлённых и лежащих деревьев напротив Викторной оранжереи – единственная в Европе (Krussmann, 1995), представляет самые старые лиственницы в Парке, растёт с первой четверти XIX в. (Фирсов, Орлова, 2008), до 1835 г. (Связева, 2005), сейчас до 18,0 м выс. Два молодых дерева: экз. № 125 – прививка Г. А. Фирсова в 1995 г., привой с рядом стоящих деревьев, на самосев Larix sibirica, пос. 2005 г. Экз. № 126 (ближнее дерево к реке Карповка) – прививка на самосев L. dahurica, пос. 2007 г. Пл.
Larix kaempferi (Lamb.) Carr. - Лиственница Кемпфера. 5 экз. Группа из трёх деревьев на уч. 57 выращивается с 1863 г. из семян от К. И. Максимовича из Японии (Связева, 2005). Уч. 90: семена из Канады (Оттава), пос. 27.08.1958 (Головач, 1980).
Уч. 23: ~85 лет, невысокое дерево под кронами других деревьев. Деревья этого вида образуют широкую крону с далеко отстоящими горизонтальными ветвями, особенно при свободной и одиночной посадке: 26,0х15,0 м (экз. на уч. 57 № 18). Пл.
Larix kaempferi (Lamb.) Carr. ‘Pendula’ - Лиственница Кемпфера ‘Пендула’, ф. плакучая, с голубоватой хвоёй. 2 экз., уч. 91, 98. Вегетативное потомство БИН, прививка Г. А. Фирсова на Larix sibirica, привой от В. И. Соловьёва из карантинной оранжереи БИН. Уч. 91: пос. 2007 г.; уч. 98: пос. 2012 г. Вег.
Larix kamtschatica (Rupr.) Carr. (L. kurilensis Mayr) - Лиственница камчатская. 6 экз. Уч. 107: происхождение самого старого экз. неизвестно, всх. 26.04.1956, пос. 19.05.1970 (Головач, 1980). Уч. 68 и 127: растение от К. Г. Ткаченко из экспедиции на остров Сахалин (западное побережье, окрест. Красногорска, 2000 г.), уч. 68 - пос. 2010 г.; уч. 127 – пос. 2007 г. Уч. 60 (3 экз.): семена из экспедиции Сада на Сахалин, посадки японцев, сделанные до Второй мировой войны в Южносахалинске, всх. 2005 г., пос. 2013 г. Пл. Одна из первых лиственниц по началу вегетации и охвоению весной. Редко встречается в культуре.
Larix kamtschatica (Rupr.) Carr. × L. kaempferi (Lamb.) Carr. - Лиственница гибридная. 5 экз. Уч. 59 (3 шт.), 94 (2 шт.). Все одного образца, семена из экспедиции Сада на Сахалин (лесные культуры у деревни Бамбучки, между Холмском и Южно-Сахалинском, сбор Г. А. Фирсова в сентябре 2004 г.). Всх. 2005 г. Уч. 94: пос. 2011 г. Уч. 59: пос. 2013 г.
Larix komarovii B. Kolesn. - Лиственница Комарова. 2 экз. Уч. 14, 99. Растение из экспедиции Сада в Приморский край (горы Сихотэ-Алинь, Лазовский район, верховья р. Милоградовка, горная тайга в ущелье у водопадов, 650 м над уровнем моря, сентябрь 1997 г.). Уч. 99: пос. 2013 г. Уч. 14: пос. 2015 г. Вег.
Larix laricina (Du Roi) K. Koch - Лиственница американская. 5 экз. В Саду известна до 1820 г. и выращивается без перерывов по настояшее время, однако сейчас представлена более молодыми растениями (Связева, 2005). В. В. Уханов (1936, с. 24) отмечал, что "в парке растет деревом до 15-16 м высоты". Два более старых дерева на уч. 57 (№ 33) и уч. 90 (№ 19) имеют возраст около 55 лет. Уч. 47 (3 шт.): семена из природы Канады (Монреаль). Всх. 1987 г., пос. 24.04.1995. Пл. В природе ареал доходит до Полярного круга на Аляске и в Канаде.
Larix lubarskii Sukaczеv (L. dahurica × L. kamtschatica × L. olgensis) - Лиственница Любарского. 2 экз., уч. 14. Экз. № 31 – более старое дерево ~45 лет, обильно и регулярно семеносит. Экз. № 38: семенное потомство БИН, второе поколение, семена с экз. № 31, всх. 2008 г., пос. 2013 г. Пл. Вид описан В. Н. Сукачёвым сравнительно недавно, в 1931 г., имеет небольшой ареал на российском Дальнем Востоке и Северо-Восточном Китае, в культуре очень редко.
Larix × maritima Sukaczev (L. dahurica × L. kamtschatica) - Лиственница приморская. 1 экз., уч. 29. Семена из природы Приморского края (Владивосток), всх. 1984 г., пос. 20.04.1995. Достигла 8,0 м выс. в 32 года. Вег. Растёт хуже лиственницы сибирской.
Larix × marschlinsii Coaz (L. kaempferi × L. decidua) Лиственница швейцарская. 2 экз. Уч. 12: семена из Франции, всх. 1954 г., пос. Б. Н. Замятнин в 1961 г. Уч. 33: семена из Литвы (Дубравская лесная опытная станция), всх. 1987 г., пос. 24.04.1995 г. Пл. (экз. на уч 12, единично).
Larix occidentalis Nutt. - Лиственница западная. 1 экз., уч. 133. Cемена из США, Вашингтон, всх. 1975 г., пос. в мае 1983 г., 18,6 м выс. в возрасте 41 год. В природе самая высокая из лиственниц Северной Америки, до 80 м выс., быстро растёт. Пл.
Larix olgensis A. Henry - Лиственница Ольгинская. 6 экз., уч. 48, 60 (2 экз.), 127, 128, 129, все одного образца. Семена из экспедиции Сада в Приморский край: Лазовский район, побережье Японского моря, устье р. Чёрная, с невысокого молодого дерева, в 1997 г., посев 21.04.1998, всх. 1.05.1998. Уч. 128: пос. 2004 г. Уч. 48: пос. 2005 г. Уч. 127 и 129: пос. 2007 г. Уч. 60: пос. 2012 г. Вид Красной книги РФ (2008). Пл.
Larix sibirica Ledeb. - Лиственница сибирская. 56 экз. на разных участках Парка. Самый распространённый вид лиственницы (рис. 1). Лучшие в аллейных посадках 1820-х гг. в регулярной части: уч. 24, 43, 43, 55, 69 и др. Уч. 57: семенное потомство с исторического дерева с уч. 81, описанного В. В. Ухановым (1940) как f. compacta Uchan. (недавно удалено как дерево угрозы), всх. 2006, пос. 2011 г. Уч. 29: № 19: подсадка молодого растения, самосев из парка, всх. 1983 г., пос. 1992 г. Пл., образует самосев. Введена в культуру Ботаническим садом БИН (Липский, Мейсснер, 1913-1915). В. В. Уханов (1936, с. 26) отмечал, что лиственница сибирская "В парке растет крупными деревьями и принадлежит к числу старейших обитателей его (самые крупные деревья посажены в 1820-30 гг."
Характеристика деревьев лиственницы в Ботаническом саду Петра Великого приводится в таблице. В графе 2 приводится номер участка (в числителе) и номер экземпляра (в знаменателе). Территория Парка-дендрария (около 16,7 га) разделена на 145 участков. Растения на каждом участке пронумерованы, с нанесением их местонахождений на планшеты. Это позволяет точно найти каждое дерево в натуре. В графе 3 возраст (лет) указан по состоянию на осень 2015 г. В графе 4 приводятся биометрические параметры для каждого дерева: высота (м), диаметр ствола на высоте груди (см) и проекция кроны (м), также состояние на осень 2015 г. (графа 5). Проекция кроны измерялась произвольно вдоль и поперёк. В случае имеющегося наклона ствола измерение проводилось в этом направлении и перпендикулярно к нему (с точность до 0,1 м). В случае выраженной асимметричности кроны измерялся наибольший её диаметр. Состояние деревьев по данным инвентаризации 1981 г. приводится в графе 6 таблицы. Растения расположены по группам, в порядке ухудшения балла состояния.

Рис. 1. Лиственница сибирская, солитерная посадка на уч. 4
Таблица. Характеристика деревьев лиственницы (Larix Mill.) в Ботаническом саду Петра Великого
Table. Specification of larch trees (Larix Mill.) at the Peter the Great Botanical Garden
| Название растений | № уч. и экз. | Возраст, лет | Биометрические параметры | Cосто-яние | Примечание |
| Larix archangelica | 33 / 28 | 11 | 1,40 м / -, 0,7 х 0,5 м | 1 | Посадка 2015 г., усыхания нет |
| Larix archangelica | 54 / 6 | 11 | 1,82 м / 1 см, 1,1 х 1,1 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix archangelica | 59 / 17 | 11 | 2,36 м / 1 см, 1,1 х 1,2 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix archangelica | 60 / 31 | 11 | 4,38 м / 4 см, 1,5 х 1,3 м | 1 | Посадка 2012 г. 2015: усыхания нет |
| Larix cajanderi | 33 / 27 | 11 | 1,80 м / -, 1,0 х 0,7 м | 1 | Посадка 2015 г.: усыхания нет |
| Larix cajanderi | 60 / 39 | ~15 | 2,15 м / 1 см, 0,7 х 0,7 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix cajanderi | 94 / 164 | ~18 | 9,4 м / 7 cм, 4,2 х 4,2 м | 1 | Посадка 2009 г. 2015: усыхания нет |
| Larix cajanderi | 132 / 147 | ~18 | 3,94 м / 3 см, 1,7 х 1,7 м | 1 | Посадка 2010 г. 2015: усыхания нет |
| Larix cajanderi | 132 / 148 | ~18 | 5,40 м / 5 cм, 2,5 х 1,9 м | 1 | Посадка 2010 г. 2015: усыхания нет |
| Larix dahurica f. fastigiata | 54 / 8 | 10 | 2,45 м / 1 cм, 0,9 х 0,9 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix dahurica f. fastigiata | 54 / 9 | 10 | 1,72 м / 1 cм, 0,8 х 0,8 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix decidua subsp. polonica | 123 / 26 | 25 | 11,9 м / 23 cм, 8,0 х 8,0 м | 1 | Посадка 2001 г. 2015: усыхания нет |
| Larix decidua subsp. polonica | 123 / 28 | 25 | 16,5 м / 32 cм, 7,8 х 8,2 м | 1 | Посадка 2002 г. 2015: усыхания нет |
| Larix decidua subsp. polonica | 123 / 29 | 25 | 13,2 м / 24 см, 7,4 х 8,0 м | 1 | Посадка 2001 г. 2015: усыхания нет |
| Larix decidua ‘Pendulina’ | 94 / 125 | ~21 | 10,6 м/ 17 cм, 3,8 х 4,7 м | 1 | Посадка 2005 г., первое пл. в 2006 г. 2015: усыхания нет |
| Larix decidua ‘Pendulina’ | 94/ 126 | ~21 | 9,0 м / 13 cм, 3,5 х 2,9 м | 1 | Посадка 2007 г. 2015: усыхания нет |
| Larix kaempferi | 57 / 18 | 153 | 20,0 / 54 см, 26,0 х 15,0 м | 1 | 1981: сушь не отмечена, хор. 2015: усыхания нет |
| Larix kaempferi ‘Pendula’ | 91 / 41 | ~16 | 1,30 м / - , 1,3 х 0,9 м | 1 | Посадка 2007 г. 2015: усыхания нет |
| Larix kaempferi ‘Pendula’ | 98 / 22 | ~9 | 2,30 м / 3 см, 0,4 х 0,7 м | 1 | Посадка 2012 г. 2015: усыхания нет |
| Larix kamtschatica | 60 / 36 | 11 | 3,95 м / 3 см, 1,7 х 1,8 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix kamtschatica | 60 / 37 | 11 | 3,30 м / 3 см, 2,6 х 1,7 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix kamtschatica | 60 / 38 | 11 | 4,16 м / 3 см, 2,4 х 2,3 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix kamtschatica | 69 / 26 | ~18 | 5,40 м / 5 м, 2,5 х 2,5 м | 1 | Посадка 2010 г. 2015: усыхания нет |
| Larix kamtschatica | 127 / 45 | ~19 | 7,1 м / 11 cм, 6,4 х 6,7 м | 1 | Посадка 2007 г. 2015: усыхания нет |
| Larix kamtschatica × L. kaempferi | 59 / 18 | 11 | 3,76 м / 2 см, 2,0 х 1,6 м | 1 | Посадка 2013 г., первое пл. в 2014 г. 2015: усыхания нет |
| Larix kamtschatica × L. kaempferi | 59 / 19 | 11 | 2,70 м / 2 см, 1,9 х 0,8 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix kamtschatica × L. kaempferi | 59 / 20 | 11 | 3,00 м / 2 см, 1,8 х 1,6 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix kamtschatica × L. kaempferi | 94 / 197 | 11 | 6,4 м / 4 cм, 3,0 х 3,4 м | 1 | Посадка 2011 г. 2015: усыхания нет |
| Larix kamtschatica × L. kaempferi | 94 / 198 | 11 | 6,1 м / 5 cм, 3,0 х 3,0 м | 1 | Посадка 2011 г. 2015: усыхания нет |
| Larix komarovii | 14 / 39 | ~22 | 2,75 м / 2 cм, 2,0 х 2,4 м | 1 | Посадка 2015 г. 2015: усыхания нет |
| Larix komarovii | 99 / 28 | ~21 | 0,92 м / -, 1,3 х 0,8 м | 1 | Посадка 2013 г. 2015: усыхания нет |
| Larix laricina | 47 / 25 | 29 | 10,0 м / 11 cм, 2,3 х 2,4 м | 1 | Посадка 1995 г. 2015: усыхания нет |
| Larix laricina | 47 / 26 | 29 | 11,2 м / 10 cм, 2,6 х 2,2 м | 1 | Посадка 1995 г., первое пл. в 2008 г. 2015: усыхания нет |
| Larix laricina | 90 / 19 | ~55 | 12,5 м / 21 cм, 5,0 х 4,5 м | 1 | 1981: сушь не отмечена, крона густая, хор. 2015: усыхания нет |
| Larix lubarskii | 14 / 31 | ~45 | 16,8 м / 28 cм, 10,0 х 10,0 м | 1 | 1981: cушь не отмечена, хор.2012: усыхания нет.2015: усыхания нет |
| Larix lubarskii | 14 / 38 | 8 | 2,03 м / 1 cм, 1,1 х 1,5 м | 1 | Посадка 2013 г.2015: усыхания нет, первое пл. в 2015 г. |
| Larix olgensis | 48 / 41 | 18 | 7,6 м / 10 cм, 4,5 х 3,9 м | 1 | Посадка 2005 г.2015: усыхания нет |
| Larix olgensis | 60 / 32 | 18 | 7,0 м / 8 см, 2,7 х 2,7 м | 1 | Посадка 2012 г.2015: усыхания нет |
| Larix olgensis | 60 / 33 | 18 | 6,6 м / 7 см, 2,5 х 3,5 м | 1 | Посадка 2012 г.2015: усыхания нет |
| Larix olgensis | 127 / 46 | 18 | 7,4 м / 14 см, 6,2 х 6,8 м | 1 | Посадка 2007 г.2015: усыхания нет |
| Larix olgensis | 128 / 3 | 18 | 9,8 м / 16 cм, 6,5 х 5,5 м | 1 | Посадка 2004 г.2015: усыхания нет |
| Larix olgensis | 129 / 8 | 18 | 7,5 м / 12 см, 5,6 х 6,0 м | 1 | Посадка 2007 г., первое пл. в 2013 г.2015: усыхания нет |
| Larix sibirica | 57 / 44 | 10 | 5,00 м / 6 cм, 3,2 х 3,0 м | 1 | Посадка 2011 г.2015: усыхания нет |
| Larix archangelica | 33 / 22 | ~200 | 30,4 м / 86 cм, 6,3 х 9,2 м | 2 | 1981: cушь не отмечена, уд. 2008: усыхание 20% кроны. 2014: усыхание 25% кроны. 2015: усыхание усилилось, 40% кроны |
| Larix archangelica | 30 / 1 | ~200 | 27,6 м / 86 cм, 10,6 х 6,3 м | 2 | 1981: ствол сильно наклонён, cушь не отмечена, уд. 2007: сушь не отмечена. 2013: отмечено усыхание, вверху заметный наклон ствола. 2015: усыхание 30% кроны |
| Larix archangelica | 29 / 2 | ~200 | 28,6 м / 71 см, 12,0 х 10,6 м | 2 | 1981: cушь не отмечена, уд. 2012: отмечена сушь. 2013: подтверждена сушь, наклон ствола вверху. 2015: усыхание 25% кроны |
| Larix × czekanowskii | 14 / 10 | ~190 | 26,0 м / 80 cм, 11,0 х 10,4 м | 2 | 1981: cушь не отмечена, хор. 2007: сушь не отмечена. 2012: усыхание незаметно, не отмечено. 2015: состояние хорошее, усыхание менее 20% кроны |
| Larix × czekanowskii | 83 / 14 | ~75 | 12,4 м / 39 cм, 6,8 х 8,5 м | 2 | 1981: крона редкая, ствол искривлён, сушь не отмечена, уд. 2008: заметно подсыхание побегов после двух последних тёплых зим. 2015: небольшое усыхание, до 20% кроны; верхушка сильно искривлена, была сломана |
| Larix × czekanowskii | 117 / 4 | ~140 | 26,8 м / 84 cм, 15,5 х 13,5 м | 2 | 1981: крона густая, низко опущена, сухая ветвь, хор. 2008: стали сохнуть ветви в кроне после недавних тёплых зим. 2015: усыхание 25% кроны |
| Larix dahurica | 14 / 13 | ~200 | 28,0 м / 60 см, 11,8 х 7,5м | 2 | 1981: cушь не отмечена, хор. 2007: сушь не отмечена. 2012: усыхание незаметно, не отмечено. 2013: состояние хорошее, сушь мало заметна. 2015: усыхание 25% кроны |
| Larix dahurica | 14 / 15 | ~200 | 29,0 м / 69 cм, 16,5 х 12,0 м | 2 | 1981: cушь не отмечена, хор. 2007: сушь не отмечена. 2012: усыхание незаметно. 2014: усыхание 15% кроны. 2015: усыхание 25% кроны |
| Larix dahurica | 14 / 16 | ~200 | 28,2 м / 72 см, 13,0 х 14,5 м | 2 | 1981: cушь не отмечена, хор. 2007: сушь не отмечена. 2012: усыхание незаметно. 2014: усыхание 20% кроны. 2015: усыхание 25% кроны |
| Larix dahurica | 14 / 17 | ~200 | 27,8 м / 71 cм, 17,0 х 10,8 м | 2 | 1981: cушь не отмечена, на выс. 8 м ствол раздвоен, хор. 2007: сушь не отмечена. 2012: усыхание незаметно. 2014: усыхание 20% кроны. 2015: усыхание 25% кроны |
| Larix dahurica | 14 / 20 | ~200 | 31,6 м / 80 cм, 13,0 х 13,2 м | 2 | 1981: cушь не отмечена, крона густая, хор. 2007: сушь не отмечена. 2012: усыхание мало заметно. 2013: сухих ветвей немного, один из лучших экз., высокая декоративность. 2015: усыхание 25% кроны |
| Larix dahurica | 14 / 21 | ~200 | 30,8 м / 86 cм, 12,0 х 11,5 м | 2 | 1981: cушь не отмечена, крона густая, хор. 2007: сушь не отмечена. 2012: усыхание мало заметно. 2014: усыхание 20% кроны. 2015: усыхание 25% кроны |
| Larix dahurica | 130 / 35 | ~160 | 27,0 м / 76 см, 7,8 х 18,0 м | 2 | 1981: крона средней густоты раскидистая, сухие ветви, хор. 2015: усыхание 25% кроны |
| Larix dahurica | 129 / 3 | ~200 | 27,0 м / 100 см, 22,2 х 12,5 м | 2 | 1981: ствол раздвоен, сушь не отмечена, хор. 2008: заметно усыхание. 2015: усыхание продолжается, после чистки сухих ветвей в кроне альпинистами зимой 2011/12 г. сохраняется высокая декоративность |
| Larix dahurica | 82 / 6 | ~200 | 27,0 м / 97 cм, 17,0 х 15,0 м | 2 | 1981: крона средней густоты, сухие ветви, уд. 2015: небольшое усыхание, до 20% кроны |
| Larix dahurica | 82 /11 | ~200 | 21,0 м / 111 см, 15,0 х 24,5 м | 2 | 1981: крона средней густоты раскидистая, сухие ветви, уд. 2015: небольшое усыхание, до 20% кроны |
| Larix decidua | 14 / 18 | ~200 | 28,4 м / 82 см, 12,0 х 13,5 м | 2 | 1981: cушь не отмечена, крона густая, хор. 2007: сушь не отмечена. 2012: усыхание незаметно. 2014: усыхание небольшое, 15% кроны. 2015: усыхание 25% кроны |
| Larix decidua | 14 / 19 | ~95 | 21,2 м / 39 cм, 8,5 х 7,5 м | 2 | 1981: крона редкая, ствол в верхней части искривлён, сухие ветви, хор. 2007: сушь не отмечена. 2012: усыхание мало заметно. 2014: усыхание 20% кроны. 2015: усыхание 25% кроны |
| Larix decidua | 32 / 21 | 32 | 4,0 м / 6 cм, 3,0 х 3,4 м | 2 | Посадка 1992 г., первое пл. в 2012 г. 2015: небольшое усыхание тонких побегов |
| Larix decidua | 48 / 23 | ~200 | 26,0 м / 113 см, 22,0 х 16,5 м | 2 | 1981: cтвол наклонён, раздвоен, сушь не отмечена, уд. 2013: отмечена небольшая сушь, до 15% кроны. 2014: усыхание до 25% кроны (немного для такого возраста, декоративность сохраняется). 2015: примерно в том же состоянии |
| Larix decidua | 48 / 24 | ~200 | 30,0 м / 101 см, 9,8 х 10,0 м | 2 | 1981: cтвол немного изогнут, сушь не отмечена, уд. 2015: усыхание 25-30% кроны, в несколько худшем состоянии по сравнению с соседним деревом |
| Larix decidua | 94 / 47 | ~85 | 25,0 м / 38 cм, 7,0 х 7,0 м | 2 | 1981: ствол сильно наклонён, крона средней густоты, односторонняя, сухие ветви, уд. 2015: усыхание небольшое, тонкие побеги |
| Larix decidua | 94 / 48 | ~85 | 22,6 м / 30 см, 4,9 х 6,9 м | 2 | 1981: ствол сильно изогнут, крона редкая, сушь не отмечена, хор. 2015: усыхание небольшое, тонкие побеги |
| Larix decidua | 145 / 54 | ~140 | 20,6 м / 54 cм, 13,5 х 14,3 м | 2 | 1981: крона редкая, односторонняя, ствол искривлён, сухие сучья, уд. 2013: заметно усыхание. 2015: усыхание 30%, крона редеет |
| Larix decidua ‘Pendula’ | 94 / 49 | ~85 | 21,2 м / 34 см, 8,0 х 8,3 м | 2 | 1981: сушь не отмечена, хор. 2015: усыхание небольшое, в основном тонкие побеги, но верхушка усыхает |
| Larix decidua ‘Pendulina’ | 94 / 2 | ~200 | 18,0 м/ 54 cм, 17,0 х 11,2 м | 2 | 1981: крона средней густоты, грибы, дупло у корня; сухие ветви, сучья, уд. 2015: усыхание небольшое |
| Larix decidua ‘Pendulina’ | 94 / 3а | ~200 | 4,10 / 49 м, 23,8 х 6,8 м | 2 | 1981: крона редкая, сухие ветви, уд. 2015: усыхание небольшое, плодовые тела трутовиков на стволе |
| Larix decidua ‘Pendulina’ | 94 / 3б | ~200 | 12,6 м / 41 cм, 9,0 х 8,5 м | 2 | 1981: крона редкая, сухие ветви, уд. 2015: усыхание небольшое, тонкие побеги |
| Larix decidua f. microcarpa | 35 / 103 | ~135 | 21,4 м / 51 см, 4,1 х 6,0 м | 2 | 1981: cушь не отмечена, хор. 2012: небольшое усыхание. 2014: усыхание 20% кроны. 2015: примерно в том же состоянии |
| Larix decidua f. microcarpa | 35 / 104 | ~ 85 | 13,2 м / 33 см, 6,9 х 6,0 м | 2 | 1981: cушь не отмечена, хор. 2012: небольшое усыхание. 2014: усыхание 20% кроны. 2015: примерно в том же состоянии |
| Larix kaempferi | 57 / 19 | 153 | 20,4 / 44 см, 16,5 х 10,5 м | 2 | 1981: сушь не отмечена, хор. 2015: сухих ветвей очень немного |
| Larix kaempferi | 57 / 20 | 153 | 21,0 м / 74 см, 21,5 х 22,6 м | 2 | 1981: сушь не отмечена, хор. 2015: сухих ветвей очень немного |
| Larix kaempferi | 90 / 17 | ~75 | 17,5 м / 47 см, 13,5 х 12,0 м | 2 | 1981: крона многовершинная, средней густоты, раскидистая, ствол слегка наклонён, сухой сук, хор. 2014: небольшое усыхание, до 15% кроны. 2015: усыхание 20% , внизу и середине кроны |
| Larix kamtschatica | 107 / 8 | 60 | 16,0 м / 40 cм, 17,0 х 12,0 м | 2 | 1981: крона редкая, сушь не отмечена, хор. 2012: сушь не отмечена. 2015: усыхание слабое, тонкие побеги |
| Larix laricina | 47 / 27 | 29 | 8,6 м / 7 cм, 2,3 х 2,2 м | 2 | Посадка 1995 г. 2015: небольшое усыхание тонких побегов |
| Larix laricina | 57 / 33 | ~55 | 23,8 м / 45 cм, 9,0 х 7,0 м | 2 | 1981: сушь не отмечена, крона густая, хор. 2008: крона рыхлая и редкая. 2013: усыхание менее 25% кроны, грибы у корневой шейки. 2015: сухих ветвей немного, в том же состоянии |
| Larix maritima | 29 / 17 | 32 | 8,0 м / 8 cм, 3,9 х 3,8 м | 2 | Посадка 1995 г. 2008: заметна сушь, засохли нижние ветви. 2012: подтверждена сушь. 2015: крона редкая, хвоя мелкая, растёт хуже соседних деревьев лиственницы сибирской |
| Larix × marschlinsii | 33 / 26 | 29 | 4,30 м / 5 см, 1,6 х 1,2 м | 2 | Посадка 1987 г. 2012: сушь не отмечена. 2015: слабое усыхание тонких побегов |
| Larix × marschlinsii | 12 / 11 | ~55 | 18,4 м / 20 cм, 3,5 х 3,0 м | 2 | 1981: cушь не отмечена, хор. 2009: усыхание не отмечено. 2012: усыхание незаметно. 2015: усыхание мало заметно, менее 15-20% кроны |
| Larix occidentalis | 133 / 31 | 41 | 18,6 м / 25 cм, 6,0 х 4,2 м | 2 | Посадка 1983 г. 2015: небольшое усыхание тонких ветвей |
| Larix sibirica | 14 / 22 | ~55 | 17,0 м / 17 cм, 5,0 х 3,5 | 2 | 1981: cушь не отмечена,крона редкая, хор. 2007: сушь не отмечена. 2012: усыхание мало заметно. 2013: отмечено усыхание, дерево угнетено, под кроной других деревьев. 2015: усыхание 25% кроны, дупло внизу, смолотечение ствола |
| Larix sibirica | 17 / 42 | ~85 | 21,4 м / 49 cм, 6,0 х 6,9 м | 2 | 1981: крона редкая, сухие ветви, уд. 2007: усыхание не отмечено. 2012: усыхание 10% кроны. 2013: подтверждено усыхание, пока немного. 2014: усыхание до 25% кроны, тонкие ветви. 2015: усыхание немного усилилось, во всех частях кроны |
| Larix sibirica | 29 / 19 | 33 | 16,0 м / 17 cм, 3,5 х 3,7 м | 2 | Посадка 1992 г. 2011: первое пл., сушь не отмечена. 2013: есть усыхание. 2015: усыхание 20% кроны |
| Larix sibirica | 32 / 17 | ~200 | 26,4 м / 81 см, 4,0 х 6,9 м | 2 | 1981: крона редкая, cушь не отмечена, уд. 2012: отмечена сушь. 2015: усыхание слабое, до 20% кроны |
| Larix sibirica | 34 / 20 | ~200 | 27,8 м / 93 cм, 12,0 х 6,5 м | 2 | 1981: cушь не отмечена, уд. 2013: отмечено небольшое усыхание, до 15% кроны. 2015: заметно сохнет, 25% кроны. 2015% усыхание 30% кроны |
| Larix sibirica | 55 / 19 | ~200 | 28,6 м / 73 cм, 9,7 х 9,0 м | 2 | 1981: ствол наклонен, крона средней густоты, сухие сучья, уд. 2013: отмечена небольшая сушь. 2015: усыхание 25% кроны |
| Larix sibirica | 60 / 28 | ~200 | 27,4 м / 83 cм, 10,0 х 16,0 м | 2 | 1981: сушь не отмечена, уд. 2010: много сухих ветвей. 2012: усыхание 20% кроны. 2013: усыхание 25% кроны. 2015: усыхание 30% кроны |
| Larix sibirica | 74 / 18 | ~190 | 23,6 м / 79 см, 16,3 х 11,8 м | 2 | 1981: Ствол раздвоен, сухие сучья, уд. 2013: усыхание 30% кроны. 2015: усыхание 35% кроны |
| Larix sibirica | 81 / 13 | ~160 | 20,6 м / 60 см, 11,0 х 10,5 м | 2 | 1981: крона густая, вершина наклонена, сухие ветки, уд. 2012: отмечена сушь. 2015: усыхание 25% кроны |
| Larix sibirica | 81 / 15 | ~200 | 23,8 м / 84 м, 16,0 х 20,0 м | 2 | 1981: крона густая раскидистая, сухие ветки, уд. 2013: усыхание 20% кроны. 2015: примерно в том же состоянии |
| Larix sibirica | 127 / 24 | ~140 | 25,5 м / 67 см, 10,0 х 11,0 м | 2 | 1981: крона средней густоты раскидистая, сухие ветви, хор. 2010: отмечено усыхание. 2013: усыхание 25% кроны. 2015: примерно в том же состоянии |
| Larix sibirica | 127 / 26 | ~140 | 22,0 м / 54 cм, 8,8 х 6,3 м | 2 | 1981: сушь не отмечена, уд. 2010: отмечено усыхание. 2013: усыхание 25% кроны. 2015: примерно в том же состоянии |
| Larix sibirica | 130 / 19 | ~150 | 25,0 м / 67 см, 7,2 х 8,4 м | 2 | 1981: ствол слегка наклонён, крона средней густоты, сухие ветви, хор. 2015: усыхание 25% , крона редеет |
| Larix sibirica | 141 / 13 | ~160 | 24,6 м / 82 см, 11,0 х 10,6 м | 2 | 1981: крона низкая, ствол изогнут у вершины, сухие сучья, уд. 2013: усыхание 20% кроны. 2015: примерно в том же состоянии |
| Larix sibirica | 31 / 6 | ~200 | 29,2 м / 93 cм, 7,2 х 7,0 м | 2 | 1981: крона редкая, дупло, cушь не отмечена, уд. 2004: плодовые тела грибов, скоро может стать деревом угрозы. 2012: отмечена сушь. 2014: усыхание 30% кроны. 2015: усыхание более 30% кроны, оголённая древесина, ствол расщеплён |
| Larix sibirica | 32 / 12 | ~200 | 29,0 м / 77 cм, 7,1 х 6,7 м | 2 | 1981: крона редкая, cушь не отмечена, уд. 2012: отмечено усыхание. 2013: усыхание небольшое, до 15% кроны. 2014: усыхание 25% кроны, плодовые тела грибов у корневой шейки |
| Larix sibirica | 39 / 3 | ~200 | 23,4 м / 84 см, 8,5 х 9,8 м | 2 | 1981: cухие ветви, крона средней густоты, гуще в сторону пруда, уд. 2012: засохли две живые ветви в середине кроны, усилилось усыхание этого и других деревьев лиственницы. 2013: усыхание 20%, тонкие ветви во всех частях кроны. 2015: усыхание 30% кроны |
| Larix sibirica | 41 / 5 | ~200 | 30,2 м / 73 см, 9,0 х 3,5 м | 2 | 1981: сушь не отмечена, уд. 2012: отмечено небольшое усыхание. 2015: усыхание до 25% кроны |
| Larix sibirica | 53 / 4 | ~200 | 27,0 м / 71 см, 7,0 х 12,0 м | 2 | 1981: крона средней густоты, сухие сучья, уд. 2008: дупло у корневой шейки. 2012: отмечены сухие ветви внизу кроны. 2014: усыхание 20% кроны. 2015: усыхание 25% кроны |
| Larix sibirica | 4 / 4 | ~125 | 24,8 м / 79 см, 16,7 х 13,6 | 2 | 1981: cушь не отмечена, хор. 2009: усыхание не отмечено. 2012: усыхание незаметно. 2013: заметно усыхание. 2014: усыхание 20% кроны. 2015: усыхание 25% кроны |
| Larix sibirica | 4 / 5 | ~125 | 23,0 м / 67 cм, 14,8 х 11,7 м | 2 | 1981: cушь не отмечена, хор. 2009: усыхание не отмечено. 2012: усыхание незаметно. 2013: заметно усыхание. 2014: усыхание 20% кроны. 2015: усыхание 25-30%, внизу и середине кроны, крона редеет |
| Larix sibirica | 24 / 5 | ~200 | 27,2 м / 85 см, 7,3 х 11,2 м | 2 | 1981: cушь не отмечена, хор. 2012: отмечено усыхание. 2013: дерево с мощным высоко очищенным от сучьев стволом, усыхание небольшое, до 20% кроны. 2015: усыхание 25% кроны |
| Larix sibirica | 24 / 6 | ~200 | 26,4 м / 81 cм, 8,5 х 8,2 м | 2 | 1981: cушь не отмечена, хор. 2012: отмечено усыхание. 2013: усыхание небольшое, до 20% кроны. 2015: усыхание 25% кроны |
| Larix sibirica | 14 / 11 | ~190 | 28,0 м / 58 см, 5,5 х 7,0 м | 2 | 1981: cушь не отмечена, хор. 2007: сушь не отмечена. 2012: усыхание незаметно, не отмечено. 2013: заметно усыхание. 2014: усыхание 20% кроны. 2015: усыхание 25% кроны |
| Larix sibirica | 14 / 12 | ~190 | 26,2 м / 74 cм, 14,0 х 9,8 м | 2 | 1981: cушь не отмечена, хор. 2007: сушь не отмечена. 2012: усыхание незаметно, не отмечено. 2013: состояние хорошее, сушь мало заметна. 2015: усыхание 25% кроны |
| Larix archangelica | 24 / 2 | ~200 | 26,2 м / 67 cм, 9,3 х 5,4 м | 3 | 1981: cухие ветки, уд. 2012: отмечено усыхание. 2013: подтверждено усыхание, плодовые тела грибов вблизи корневой шейки. 2014: усыхание 25% кроны. 2015: усыхание 30% кроны |
| Larix archangelica | 29 / 14 | ~200 | 29,0 / 85 cм, 8,5 х 9,2 м | 3 | 1981: cушь не отмечена, уд. 2007: сушь не отмечена. 2012: отмечена сушь. 2013: усыхание стало очень заметным. 2014: усыхание до 30% кроны. 2015: усыхание более 30% кроны |
| Larix dahurica | 27/ 5 | ~200 | 29,2 м / 113 см, 17,5 х 18,0 м | 3 | 1981: две вершины, cухие сучья, уд. 2013: давно заметна сушь. 2015: усыхание 35% кроны |
| Larix dahurica | 27 / 6 | ~200 | 20,2 м / 95 cм, 11,5 х 10,5 м | 3 | 1981: сухие сучья, капы, уд. 2009: сломан ствол, есть сушь. 2013: усыхание усилилось. 2015: усыхание 35% кроны |
| Larix dahurica | 58 / 12 | ~200 | 29,2 м / 94 см, 8,5 х 21,5 м | 3 | 1981: крона редкая, сухие ветви, уд. 2012: отмечено усыхание. 2013: усыхание 30% кроны, грибы у корневой шейки. 2015: усыхание 35% кроны |
| Larix dahurica | 58 / 13 | ~200 | 30,0 м / 63 cм, 7,8 х 11,5 м | 3 | 1981: сушь не отмечена, хор. 2013: усыхание 20% кроны. 2015: усыхание усилилось, 35% кроны |
| Larix decidua | 14 / 14 | ~200 | 28,4 м / 100 cм, 19,0 х 17,0 м | 3 | 1981: cушь не отмечена, хор. 2007: сушь не отмечена. 2012: усыхание незаметно. 2013: состояние хорошее, но есть сухие ветви. 2014: усыхание 25% кроны. 2015: усыхание 30% кроны |
| Larix decidua | 92 / 23 | ~160 | 26,0 м / 68 cм, 11,8 м х 6,5 м | 3 | 1981: крона густая раскидистая, сухие сучья, уд. 2008: отмечено усыхание. 2013: крона редеет, усыхание 15% кроны. 2015: усыхание усилилось, 35%, сухие ветви по всей кроне |
| Larix kaempferi | 23 / 32 | ~85 | 7,4 м / 28 cм, 8,7 х 8,3 м | 3 | 1981: крона средней густоты раскидистая, ствол слегка наклонён, cушь не отмечена, хор. 2012: cушь не отмечена. 2013: отмечено усыхание. 2015: усыхание около 30% кроны, верхушка параллельна поверхности почвы, под пологом деревьев дуба черешчатого |
| Larix sibirica | 3 / 50 | ~115 | 24,2 м / 47 см, 4,2 х 2,5 м | 3 | 1981: ствол слегка искривлён, крона средней густоты, сухие ветки, уд. 2007: усыхание 25% кроны, сырое место на границе затопляемого участка. 2014: 40% кроны. 2015: 50% кроны |
| Larix sibirica | 3 / 51 | ~115 | 25,6 м / 61 cм, 7,4 х 6,7 м | 3 | 1981: сухие ветки, уд. 2007: сохнет, 25% кроны. 2014: усыхание 30% кроны. 2015: примерно в том же состоянии |
| Larix sibirica | 14 / 9 | ~200 | 25,0 м / 67 cм, 10,6 х 8,5 м | 3 | 1981: cушь не отмечена, хор. 2007: сушь не отмечена. 2012: много сухих ветвей. 2013: сохнет, до 35% кроны. 2014: усыхание усилилось, 40% кроны. 2015: усыхание более 40% кроны |
| Larix sibirica | 24 / 7 | ~200 | 26,0 м / 76 cм, 13,4 х 11,4 м | 3 | 1981: крона односторонняя, ствол слегка наклонен, сухие ветви, уд. 2012: отмечено усыхание. 2013: подтверждено усыхание. 2015: заметно хуже других деревьев этого участка, сухие скелетные ветви |
| Larix sibirica | 25 / 8 | ~200 | 26,4 м / 83 см, 12,5 х 16,4 м | 3 | 1981: cухие сучья, уд. 2012: отмечено усыхание. 2014: усыхание 30% кроны. 2015: примерно в том же состоянии |
| Larix sibirica | 31 / 26 | ~200 | 27,8 м / 89 см, 7,0 х 10,0 м | 3 | 1981: крона редкая, cушь не отмечена, уд. 2012: усыхание 15-20% кроны. 2014: усыхание 25% кроны, плодовые тела грибов у корневой шейки. 2015: усыхание 30% кроны |
| Larix sibirica | 37 / 36 | ~200 | 27,4 м / 88 cм, 16,0 х 13,0 м | 3 | 1981: cухие сучья, крона средней густоты, уд. 2012: усыхание 20% кроны. 2013: усыхание 25% кроны. 2014: усыхание 30% кроны. 2015: усыхание 35% кроны, заметный наклон ствола, особенно в верхней части кроны |
| Larix sibirica | 42 / 18 | ~200 | 28,6 м / 90 cм, 8,2 х 10,0 м | 3 | 1981: сушь не отмечена, уд. 2012: появилась сушь, 20% ветвей. 2014: усыхание усилилось, 30% кроны. 2015: усыхание 40% кроны |
| Larix sibirica | 42 / 34а | ~200 | 30,6 м / 66 cм, 5,5 х 8,0 м | 3 | 1981: сушь не отмечена, уд. 2012: усыхание 20% кроны. 2013: усыхание 30% кроны. 2014: усыхание усилилось, 40% кроны |
| Larix sibirica | 43 / 27 | ~200 | 27,2 м / 57 cм, 5,2 х 3,5 м | 3 | 1981: крона средней густоты на верхушке, сухие ветки, уд. 2012: усыхание 25% кроны. 2014: усыхание 30% кроны. 2015: примерно в том же состоянии |
| Larix sibirica | 46 / 10 | ~200 | 26,4 м / 88 см, 11,1 х 12,5 м | 3 | 1981: крона редкая, ствол наклонён, сухие ветки, уд. 2012: есть усыхание. 2013: усыхание 25% кроны, грибы у шейки корня. 2015: усыхание 30% кроны |
| Larix sibirica | 46 / 20 | ~200 | 26,0 м / 87 см, 9,2 х 6,9 м | 3 | 1981: ствол наклонён, сухие ветви, уд. 2008: отмечена сушь, крона редкая и маленькая. 2012: усыхание 30% кроны. 2015: усыхание 40% кроны |
| Larix sibirica | 52 / 24 | ~200 | 26,4 м / 67 см, 8,7 х 7,8 м | 3 | 1981: крона редкая, сушь не отмечена, уд. 2007: крона редкая, есть усыхание. 2012: усыхание 25% кроны. 2013: усыхание 35% кроны. 2015: усыхание 50% кроны |
| Larix sibirica | 54 / 4 | ~200 | 28,4 м / 85 см, 8,0 х 10,3 м | 3 | 1981: ствол слегка наклонён, крона средней густоты, сухие сучья, уд. 2013: усыхание 25% кроны. 2015: заметно сохнет, 35% кроны |
| Larix sibirica | 55 / 2 | ~200 | 26,8 м / 70 cм, 12,2 х 7,5 м | 3 | 1981: крона односторонняя средней густоты, сухие ветки, уд. 2013: усыхание 30% кроны. 2015: усыхание 35%, крона редеет |
| Larix sibirica | 55 / 9 | ~200 | 28,4 м / 78 cм, 9,5 х 10,5 м | 3 | 1981: крона односторонняя средней густоты, сухие ветки, уд. 2013: усыхание 20% кроны. 2014: усыхание 25% кроны. 2015: усыхание 30% кроны |
| Larix sibirica | 55 / 18 | ~200 | 28,6 м / 82 cм, 10,8 х 7,0 м | 3 | 1981: крона односторонняя средней густоты, сухие сучья, уд. 2013: усыхание 20% кроны. 2015: усыхание 30% кроны. Плодовые тела грибов у корневой шейки дерева |
| Larix sibirica | 55 / 54 | ~135 | 22,0 м / 55 см, 10,5 х 10,2 м | 3 | 1981: крона односторонняя средней густоты, сухие сучья, уд. 2012: есть усыхание. 2013: усыхание 25% кроны, грибы у корневой шейки. 2014: усыхание 30% кроны. 2015: усыхание 35% кроны |
| Larix sibirica | 57 / 7 | ~140 | 23,2 м/ 54 см, 10,0 х 8,0 м | 3 | 1981: крона средней густоты, сухие ветви, хор. 2012: усыхание 20% внизу и середине кроны. 2013: усыхание 30% кроны. 2014: усыхание усиливается, 40% кроны. 2015: усыхание 60% кроны |
| Larix sibirica | 58 / 14 | ~200 | 29,0 м / 70 cм, 7,5 х 7,2 м | 3 | 1981: сушь не отмечена, хор. 2012: отмечено усыхание. 2013: усыхание 20% кроны. 2014: усыхание 25% кроны. 2015: усыхание 30% кроны. |
| Larix sibirica | 59 / 7 | ~200 | 30,0 м / 73 cм, 10,0 х 8,3 м | 3 | 1981: сушь не отмечена, хор. 2013: усыхание 30% кроны. 2015: усыхание 35% кроны |
| Larix sibirica | 59 / 11 | ~200 | 29,4 м / 96 cм, 9,2 х 14,0 м | 3 | 1981: крона средней густоты, сухие ветки, дупло у корня, уд. 2008: дупло, крона редкая, есть сухие ветви. 2013: усыхание 25% кроны. 2014: усыхание 35% кроны. 2015: усыхание 40% кроны, засохли скелетные ветви во всех частях кроны |
| Larix sibirica | 69 / 24 | ~200 | 28,6 м / 72 cм, 10,0 х 9,5 м | 3 | 1981: ствол слегка наклонён, крона редкая, сухие ветви, уд. 2012: отмечено усыхание. 2013: усыхание 30% кроны. 2015: усыхание 35% кроны |
| Larix sibirica | 89 / 32 | ~100 | 18,0 м / 42 м, 8,5 х 8,5 м | 3 | 1981: крона средней густоты, относторонняя, сухие сучья, уд. 2008: отмечено подсыхание концов побегов. 2013: усыхание 30% кроны. 2015: примерно в том же состоянии |
| Larix sibirica | 117 / 29 | ~140 | 20,4 м / 56 см, 11,0 х 10,8 м | 3 | 1981: крона средней густоты раскидистая, ствол немного наклонён, вершина опилена, сухие ветви, хор. 2008: стали сохнуть ветви в кроне. 2012: усыхание 25% кроны. 2014: усыхание 30% кроны. 2015: усыхание 35% кроны |
| Larix sibirica | 126 / 37 | ~160 | 25,6 м / 73 cм, 8,0 х 8,5 м | 3 | 1981: крона средней густоты, сухие ветви, уд. 2008: много сухих ветвей. 2013: усыхание 30% кроны. 2015: усыхание 35% кроны |
| Larix sibirica | 128 / 50 | ~140 | 23,6 м / 59 см, 9,0 х 10,0 м | 3 | 1981: две вершины, крона односторонняя редкая, сухие сучья, уд.2013: усыхание 20% кроны. 2015: усыхание заметно, 30% кроны |
| Larix decidua | 140 / 47 | ~140 | 17,2 м / 54 cм, 12,0 х 10,0 м | 4 | 1981: крона редкая, сушь не отмечена, уд. 2008: много сухих ветвей после двух последних тёплых зим. 2010: наполовину сухая. 2012: усыхание 60% кроны. 2013: усыхание 70% кроны. 2014: усыхание 80% кроны. 2015: усыхание 90% кроны |
| Larix sibirica | 32 / 1 | ~200 | 28,8 м / 61 cм, 4,2 х 6,0 м | 4 | 1981: крона редкая, cушь не отмечена, уд. 2012: заметно сохнет. К 2013 г. постепенно засохло около половины кроны. 2014: усыхание 60% кроны. 2015: усыхание 70% кроны |
| Larix sibirica | 42 / 34б | ~200 | 28,6 м / 83 см, 6,5 х 16,9 м | 4 | 1981: сушь не отмечена, уд. 2012: усыхание 20% кроны. 2013: усыхание 30% кроны. 2014: усыхание 40% кроны. 2015: усыхание 60% кроны |
| Larix sibirica | 128 / 24 | ~200 | 23,2 м / 69 см, 7,6 х 10,0 м | 4 | 1981: крона средней густоты, сухие ветви, уд. 2008: много сухих ветвей после аномально тёплой зимы 2006/07 г. 2013: усыхание 40% кроны. 2015: усыхание 50% кроны (самый плохой экз. после L. decidua на уч. 140) |
Всего в таблицу включены 148 экземпляров лиственницы (Larix Mill.), относящихся к 22 видам и формам, которые выращиваются в Ботаническом саду Петра Великого БИН в Санкт-Петербурге. Преобладает L. sibirica (56 экз.), за которой следуют, намного уступая ей в количестве экземпляров, L. dahurica (14) и L. decidua (11 экз.). Остальные таксоны составляют меньшинство, от 9 экз. (L. archangelica) и менее, при этом L. decidua f. pendula, L. × maritima и L. occidentalis представлены в единственном числе.
Представители рода Larix относятся к наиболее долговечным древесным растениям в Саду. Многие деревья лиственницы сохранились с самых первых посадок, сделанных в Саду в 1820-х гг. после преобразования бывшего Аптекарского огорода в Императорский Санкт-Петербургский Ботанический сад. Если рассмотреть распределение деревьев лиственницы по классам возраста (если принять класс возраста равным 10 лет), то к молодым, I-III классов возраста, можно отнести 42 экз. Наличие такого количества молодых особей (28%) свидетельствует об интенсивной интродукционной работе с коллекцией в последние десятилетия. Лиственница относится к быстрорастущим древесным породам, 5 особей – первого класса возраста (до 11 лет), уже в таком возрасте деревья могут высаживаться из питомника на постоянное место в парк. Насчитывается 20 деревьев, которые можно отнести к «приспевающим», от IV до X классов возраста (до 100 лет). Старых деревьев, старше 100 лет – 86 экз., то есть большинство. При этом наибольшее их число, 59 экз. или 38% от общего числа, можно отнести к историческим, посаженным в 1820-е годы, после преобразования в 1823 г. бывшего Аптекарского огорода в Императорский Санкт-Петербургский Ботанический Сад, когда были заложены аллеи. Именно лиственница (преимущественно лиственница сибирская, с участием лиственниц даурской, архангельской и европейской) образует основу древостоя парка-дендрария, она же вместе с дубом черешчатым, липой сердцевидной и клёном остролистным является видом-образователем аллейных посадок в регулярной части парка. По продолжительности жизни с лиственницей может сравниться только дуб черешчатый (Quercus robur L.), деревья которого достигают такого же значительного возраста, как и виды рода Larix. Однако в последние годы состояние дуба, как и лиственницы, также заметно ухудшилось (Веденяпина и др., 2015). Клён остролистный (Acer platanoides L.) в аллейных посадках представлен заметно более молодыми деревьями, возраста до 100-120 лет – он менее долговечен, а вязы (Ulmus laevis Pall. и др.) вообще почти все выпали из-за голландской болезни вязов. В последние годы и десятилетия интродукции лиственниц уделялось внимание. Из экспедиций Сада привезены и уже высажены на постоянное место в парк L. olgensis и L. komarovii – новые виды для коллекции. Особенно остро вопрос с новыми посадками лиственницы стоит в связи с необходимостью восстановления аллей в регулярной части парка, где в последние годы выпал целый ряд старых деревьев и появились прогалины, проплешины, открытые места. Как видно из таблицы, на участках 33, 54, 59, 60 в 2012-2015 гг. было высажено более 10 молодых деревьев разных видов лиственницы, некоторые из них уже стали давать шишки.
Распределение деревьев по типам посадки приводится на рис. 2. По числу особей преоблают деревья, высаженные в аллеях – 62 экз. (42%). За ними следуют солитерные посадки – 52 экз. (35%). Остальные 34 экз. представляют собой групповые посадки. Большую куртину составляет группа лиственниц из 16 экз. на уч. 14, на склонах и у подножья холма с беседкой; остальные группы из небольшого числа деревьев. Гистограмма (рис. 1) отображает количественное распределение (ось ординат) экземпляров рода Larix по категориям (ось абсцисс) состояния в каждом типе посадок. При этом прямой зависимости категории состояния от типа посадок не наблюдается.
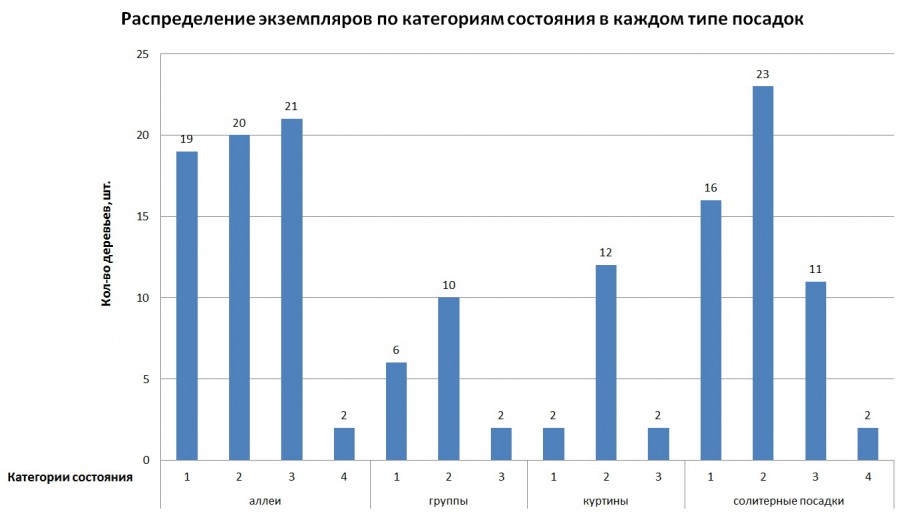
Рис. 2. Категории состояния в типе посадок.
Что касается размеров, то все виды лиственницы относятся к деревьям первой величины (по классификации С. Я. Соколова, О. А. Связевой, 1965) – группа Д1, более 25 м выс.
Некоторые деревья не достигли пока что крупных размеров из-за молодого возраста, они продолжают увеличиваться в размерах. В нескольких случаях размеры в высоту растений современной коллекции Ботанического сада Петра Великого превосходят таковые, известные в Санкт-Петербурге ранее (Булыгин и др., 1989), как например, для L. decidua subsp. polonica. Так, в настоящее время представители L. kamtschatica достигают 16,0 м выс., ранее известное значение – 12,0 м. Для таких видов, как L. komarovii, биометрические показатели в Санкт-Петербурге были неизвестны и ранее не приводились. Сейчас высоты 30 м достигают и превосходят 8 деревьев: L. dahurica (уч. 14 № 20) – 31,6 м (самое высокое дерево в коллекции); L. dahurica (уч. 14 № 21) – 30,8 м; L. sibirica (уч. 42 № 34а) – 30,6 м; L. archangelica (уч. 33 № 22) – 30, 4 м; L. sibirica (уч. 41 № 5) – 30,2 м; L. dahurica (уч. 58 № 13) – 30,0 м; L. decidua (уч. 48 № 24) – 30,0 м; L. sibirica (уч. 50 № 7) – 30,0 м. По диаметру ствола самых крупных значений достигают L. decidua (уч. 48 № 23) и L. dahurica (уч. 27 № 5) – 113 см. Сравнение с данными 1981 г. показывает, что у старых деревьев за прошедшие 35 лет в основном произошло небольшое увеличение прироста по высоте и диаметру. Так, например, L. dahurica (уч. 14 № 13, возраст сейчас ~200 лет): высота изменилась с 25,0 до 28,0 м, диаметр ствола увеличился лишь с 54 до 60 см. У некоторых деревьев изменений в размерах почти не произошло – в ряде случаев это можно объяснить наклоном ствола или засыханием верхушки, иногда обламыванием ствола. У ряда старых деревьев к этому возрасту прирост по высоте прекращается. У более молодых деревьев изменение в размерах более значительное. Так, у L. decidua (уч. 94 № 48, возраст ~85 лет) высота увеличилась с 15,0 м до 22,6 м, а диаметр ствола – с 22 см до 30 см.
По данным инвентаризации 1981 г. из сохранившихся к настоящему времени деревьев лиственницы в то время из 103 экз. - 42 дерева были оценены как в «хорошем состоянии» (41%), и 61 дерево – в «удовлетворительном» (59%), в «неудовлетворительном» не было ни одного. Об усыхании и изреживании крон деревьев в связи с фитофторами в то время не было речи. 45 деревьев современной коллекции представлены более молодыми особями и были посажены после 1981 г., поэтому тогда не могли быть учтены.
По данным современной инвентаризации к настоящему времени к первой категории состояния можно отнести 43 экз., ко второй - 65, к третьей – 34, к четвёртой – 4. Состояние и степень усыхания заметно связаны с возрастом. Это касается всех видов. При этом наблюдается некоторая тенденция, что такие виды, как L. lubarskii и L. kaempferi выглядят более устойчивыми по сравнению с другими лиственницами. Относительно слабое усыхание показывает также L. czekanowskii. К более сильно усыхающим видам можно отнести L. decidua. Что касается возраста, то к первой категории состояния относятся почти все молодые растения, от 8 лет (L. lubarskii). Они преимущественно относятся ко II-III классам возраста (если принять класс возраста равным 10 лет). Лишь в двух случаях это деревья V-VI классов возраста (L. lubarskii и L. laricina) и 1 дерево (L. kaempferi) – ещё более старшего возраста, старше 100 лет. В противоположность им, виды 3-4 категории состояния представлены наиболее старыми деревьями. Почти все они значительного возраста, до 200 лет, лишь в редких случаях моложе. Вторая категория промежуточная, она же наиболее представительная по числу особей. Там много старых деревьев, пока что ещё мало потерявших декоративность из-за усыхания. Но есть и сравнительно молодые растения. Это L. decidua (уч. 32 № 21) и L. maritima (уч. 29 № 17) – 32 года, L. sibirica (уч. 29 № 19) – 33 года, L. occidentalis (уч. 133 № 41) – 41 год. У старых деревьев вблизи корневой шейки, на корневых лапах часто можно обнаружить плодовые тела трутовика Швейница (Phaeolus schweinitzii (Fr.) Pat.) – патогена, который паразитирует на корнях хвойных представителей разных родов и вызывает бурую гниль.
При мониторинге коллекционных древесных растений парка-дендрария в 1990-е и начале 2000-х гг. куратором коллекции прежде всего обращалось внимание на повреждения побегов после каждой зимы от морозов, поскольку зимостойкость является основным фактором, ограничивающим возможность выращивания деревьев и кустарников в открытом грунте, как показал весь опыт их разведения в Санкт-Петербурге за три века интродукции (Фальк, 1766; Фирсов, Фадеева, 2009; Фирсов, 2014). Однако в начале XXI века, после аномально тёплой зимы 2006/07 гг. стало наблюдаться заметное усыхание деревьев многих видов (Фирсов и др., 2008, 2010), в том числе таких вполне и сравнительно зимостойких, как представители рода Larix, которые ранее не обмерзали даже в самые неблагоприятные аномально холодные зимы. Это заметно по данным графы 6 в таблице.
Результаты исследования ризосферной почвы лиственниц на наличие оомицетов из рода Phytophthora
Результаты проведенных исследований 2011-2014 гг. показали, что в почве парка-дендрария БИН в ризосфере различных растений распространены популяции пяти видов фитофтор. Это почвообитающие корнепоражающие виды, представляющие опасность для произрастающих в парке древесных растений: Phytophthora cactorum (Lebert et Cohn) J. Schröt., Ph. citricola Sawada, Ph. plurivora T. Jung et T. I. Burgess, Ph. quercina T. Jung и Ph. cinnamomi Rands (Веденяпина и др., 2014а,б). В образцах, которые были взяты непосредственно в ризосфере лиственниц, была обнаружена Phytophtora cactorum (Фитофтора кактусов) – исключительно под Larix decidua. Позже этот экземпляр лиственницы европейской окончательно засох и к настоящему времени удалён.

Рис. 3. Усыхание Larix decidua уч. 140.
Вид Ph. cactorum был впервые описан из Германии (как Peronospora cactorum Lebert et Cohn) как паразит кактусов. В настоящее время известно о его распространении по всему земному шару (вид является космополитным), и круг растений-хозяев этого вида постоянно увеличивается. Несмотря на свое название, Ph. cactorum известен как возбудитель фитофторозов около 150 родов сосудистых растений из более чем 50 семейств (Waterhouse, Waterston, 1966; Erwin, Ribeiro, 1996). С определенной регулярностью данный патоген идентифицируется в почвенных пробах в естественных лесных сообществах и искусственных лесопосадках, являясь возможной причиной усыхания различных древесных пород (Brasier, 2000). Причем он может присутствовать в активном состоянии в ризосфере деревьев, не имеющих никаких симптомов заболевания и вызывать быструю и бессимптомную гибель растения. Но чаще он приводит к видимым поражениям различных органов растения, не имея четкой приуроченности к локализации в тканях хозяина (может вызывать трещины коры, гниль корней, листьев и плодов). Это раневый паразит, образующий сильные токсины. Прорастание покоящихся структур (ооспор), переживающих зимний период в опавших растительных остатках, происходит весной при достижении температуры почвы 7.5° С, и затем, при постепенном прогревании почвы, продуцирование зооспорангиев с зооспорами лавинообразно нарастает, что приводит к захвату патогеном больших территорий и быстрому его распространению. Увеличение продолжительности вегетационного сезона в Санкт-Петербурге и повышение среднегодовой температуры способствует процветанию данного вида на занятой им территории.
В результате изучения почвенных проб с применением метода приманок в 2015 г. в ризосфере усыхающего дерева Larix decidua (рис. 3) был обнаружен еще один вид фитофтор. Амплификация фрагмента мДНК (COX2) и сравнение полученных последовательностей с имеющимися в международной базе данных GenBank с помощью алгоритма BlastN показало достоверное сходство наших образцов с Phytophthora syringae (Berk.) Kleb. (Фитофтора сирени). Этот вид ранее не отмечался в парке-дендрарии Ботанического сада БИН. Исследование почвы под другими видами лиственницы, как и ранее, показало отсутствие фитофтор, тогда как выявленный вид Ph. syringae доминировал в большинстве почвенных образцов многих древесных пород и кустарников.
Вид Ph. syringae был описан в конце XIX столетия как патоген, поражающий листья сирени (Syringa vulgaris L.), а его статус в роде Phytophthora утвердился лишь в 1909 году. Данный вид является близкородственным таксоном Ph. cactorum. До недавнего времени он был известен в основном как патоген деревьев из семейства Rosaceae, произрастающих в умеренном климате (Erwin, Ribeiro, 1996). Однако на сегодня накоплены данные о его паразитической активности в отношении 29 родов деревьев из 14 семейств, включая хвойные породы (Pinus и Juniperus), и распространении практически на всех континентах (Phytophthora Database, 2015). Поэтому мы не исключаем возможность его паразитической активности по отношению к видам рода Larix. Как и близкий вид, Ph. cactorum, вызывает похожие симптомы поражения растений и может приводить к ожогу ветвей, корневой гнили и гнили плодов, изъязвлению основания ствола, пятнистости листьев и появлению ложной мучнистой росы. Минимальная температура роста, необходимая для образования инфекционных структур, менее 5° С и оптимальная колеблется в пределах 15-20° С, что позволяет данному виду фитофторы сохраняться в почве в активном состоянии даже в позднеосенний период (в условиях теплой осени) (Ho, Jong, 1993).
Одной из возможных причин ухудшения состоянии лиственницы в парке-дендрарии БИН является воздействие фитофтор. Результатом проведенных исследований является достоверное выявление в парке-дендрарии Ботанического сада Петра Великого БИН корневых патогенов – видов рода Phytophthora из группы грибоподобных оомицетов, которые предположительно являются инициирующими биотическими факторами (по Manion, 1991), способными активно поражать древесные растения и кустарники, вызывая корневую гниль и усыхание растения. Паразитическая активность фитофтор в большой степени определяется внешними факторами среды, поэтому изменение климатических показателей, главным образом температуры и влажности, в сторону создания неблагоприятных для растения условий приводит к ослаблению защитной системы растений и усилению патогенных свойств видов Phytophthora. Распространение инфекции при данных условиях может быть очень быстрым и охватывать довольно большие территории.
Последние новейшие исследования показали комплексное участие многих патогенов из группы грибоподобных оомицетов в деградации лесных сообществ. Прогресс в применении новых молекулярных методов детекции и идентификации таксонов позволил установить тот факт, что наряду с представителями рода Phytophthora, компонентами комплекса активных патогенов могут выступать также виды рода Pythium (Jiménez et al., 2008; Robideau et al., 2011), ранее считавшиеся сапротрофами или слабыми патогенами. При этом общее разнообразие агрессивных агентов из фитофтор и питиумов в почвах естественных лесонасаждений может достигать 10 и более видов, присутствующих как в здоровых, так и в усыхающих древостоях (Jung et al., 2000). В условиях города и искусственных насаждений подобная картина может еще более усложняться на фоне ослабления иммунитета растений к стрессовым факторам. Кроме того, в парке-дендрарии многие интродуцированные деревья и кустарники из различных таксономических групп, в природе произрастающие в различных географически удаленных районах, соседствуют друг с другом, вынужденно разделяя общую почвенную среду. В этих условиях можно прогнозировать также изменение поведения почвенных патогенов, способных приспосабливаться к новым абиотическим и биотическим факторам, и даже менять растения-хозяев. Так, выявленный нами в ризосфере лиственницы европейской патоген Phytophthora syringae, ранее не отмечался как паразит видов рода Larix. Участие этого вида фитофторы в усыхании лиственниц, а также присутствие в ризосфере других древесных пород и кустарников парка-дендрария Ботанического сада (дуба, яблони, жимолости и рододендронов) может свидетельствовать о расширении его круга хозяев и изменении стратегии поведения именно в данных конкретных условиях.
Заключение
В Ботаническом саду Петра Великого Ботанического института им. В. Л. Комарова РАН в Санкт-Петербурге выращивается 148 экземпляров лиственницы (Larix Mill.), которые относятся к 22 видам и формам. Это долговечные, до 200-летнего возраста, декоративные деревья крупных размеров, они составляют основу древостоя парка-дендрария и образуют аллеи в наиболее старой, регулярной части парка. Самые крупные деревья по высоте достигают 31,6 м (L. dahurica), по диаметру ствола – 113 cм (L. decidua и L. dahurica).
С потеплением климата Санкт-Петербурга в начале XXI века и после аномально тёплой зимы 2006/07 гг. (Фирсов и др., 2008, 2010) и последующих тёплых зим стало наблюдаться заметное усыхание деревьев многих видов, в том числе таких вполне и сравнительно зимостойких, как представители рода Larix, которые ранее не повреждались даже в самые неблагоприятные аномально холодные зимы. Усыхание, очевидно, связано с возрастом растений и заметно усиливается у более старых деревьев. По данным современной инвентаризации к первой категории состояния можно отнести 43 экз., у которых усыхание кроны отсутствует. У остальных 105 экземпляров усыхание наблюдается в той или иной степени.
Одной из возможных причин ухудшения состоянии лиственницы в парке-дендрарии БИН является воздействие фитофтор. Результатом проведенных исследований является достоверное выявление в парке-дендрарии Ботанического сада Петра Великого БИН корневых патогенов – видов рода Phytophthora из группы грибоподобных оомицетов, которые предположительно являются инициирующими биотическими факторами, способными активно поражать древесные растения и кустарники, вызывая корневую гниль и усыхание растения.
Результаты проведенных исследований 2012-2015 гг. показали, что в почве парка-дендрария БИН, в ризосфере различных растений распространены популяции шести видов фитофтор. Это почвообитающие корнепоражающие виды, представляющие опасность для произрастающих в парке древесных растений: Phytophthora cactorum, Ph. citricola, Ph. plurivora, Ph. quercina и Ph. cinnamomi (Веденяпина и др., 2014а,б). В образцах, которые были взяты непосредственно в ризосфере лиственниц, были обнаружены Phytophtora cactorum и Ph. syringae. Последний обнаружен в Ботаническом саду БИН впервые.
Необходимо продолжение обследования насаждений лиственницы и других древесных пород парка-дендрария Ботанического сада БИН с целью выявления всех почвообитающих патогенов, способных вызвать усыхание и гибель растений. Поэтому изучение их распространения в почве, включающее детекцию, идентификацию и количественный учет, а также изучение особенностей их жизненного цикла в конкретной экологической ситуации, особенно в городских условиях, весьма актуально.
Лиственницы по-прежнему являются перспективными для разведения. Однако кроме зимостойкости, надо уделять внимание их устойчивости к фитофторам, как и другим возможным патогенам, особенно если потепление климата будет продолжаться.
Благодарности
Работа выполнена в рамках выполнения государственного задания согласно тематическому плану Ботанического института им. В. Л. Комарова РАН по теме № 0126-2014-0021. Коллекции живых растений Ботанического института им. В. Л. Комарова РАН (история, современное состояние, перспективы развития и использования).
The present study was carried out within the framework of the institutional research project (№ 0126-2014-0021) of the Komarov Botanical Institute of the Russian Academy of Sciences.
Литература
Булыгин Н. Е. Фенологические наблюдения над древесными растениями. Л.: ЛТА, 1979. 97 с.
Булыгин Н. Е. Биологические основы дендрофенологии. Л.: ЛТА, 1982. 80 с.
Булыгин Н. Е., Фирсов Г. А., Комарова В. Н. Основные результаты и перспективы дальнейшей интродукции хвойных на Северо-Западе России. Ленингр. лесотехн. акад. Деп. в ВИНИТИ 15.06.1989. № 3983—В89.
Веденяпина Е. Г., Волчанская А. В., Малышева В. Ф., Малышева Е. Ф., Фирсов Г. А. Почвообитающие виды рода Phytophtora в Ботаническом саду БИН РАН. I. Первые находки Ph. citricola, Ph. plurivora и Ph. quercina в России // Микология и фитопатология. 2014а. Т. 48. Вып. 4. С. 263—273.
Веденяпина Е. Г., Фирсов Г. А., Волчанская А. В., Воробьев Н. И. Почвообитающие виды рода Phytophthora в Ботаническом саду БИН РАН. II. Результаты двухлетнего мониторинга // Микология и фитопатология. 2014б. Т. 48. Вып. 5. С.322—332.
Веденяпина Е. Г., Фирсов Г. А. Невидимые фитофторы // Питомник и частный сад. № 2. 2014. С. 40—45.
Веденяпина Е. Г., Волчанская А. В., Лаврентьев Н. В., Фирсов Г. А. Состояние дуба черешчатого (Quercus robur L.) в Ботаническом саду БИН РАН // Вестник Удмуртского университета. Серия Биология. Науки о Земле. 2015. Т. 25. Вып. 2. С. 43—50.
Головач А. Г. Деревья, кустарники и лианы Ботанического сада БИН АН СССР (итоги интродукции). Л.: Наука, 1980. 188 с.
Красная книга Российской Федерации (растения и грибы) / Под ред. Ю. П. Трутнева и др. Сост. Р. В. Камелин и др. М.: Товарищество научных изданий КМК, 2008. 855 с.
Лапин П. И. Сезоннный ритм развития древесных растений и его значение для интродукции // Бюл. Глав. Ботан. сада. 1967. Вып. 65. С. 13—18.
Липский В. И. Исторический очерк Императорскаго С.-Петербургскаго Ботаническаго Сада // Императорский С.-Петербургский Ботанический Сад за 200 лет его существования (1713-1913). Ч. 1. СПб., 1913. 412 с.
Липский В. И., Мейсснер К. К. Перечень растений, распространенных в культуре Императорским С.-Петербургским Ботаническим садом // Императорский С.-Петербургский Ботанический сад за 200 лет его существования (1713-1913). Ч. 3. Петроград, 1913-1915. С. 537—560 с.
Мозолевская Е. Г., Катаев О. А., Соколова Э. С. Методы лесопатологического обследования очагов стволовых вредителей и болезней леса. М.: Лесная промышленность, 1984. 152 с.
Связева О. А. Деревья, кустарники и лианы парка Ботанического сада Ботанического института им. В. Л. Комарова (К истории введения в культуру). СПб.: Росток, 2005. 384 с.
Соколов С. Я., Связева О. А. География древесных растений СССР. М., Л.: Наука, 1965. 265 с.
Уханов В. В. Парк Ботанического института АН СССР. М., Л.: Изд-во АН СССР, 1936. 168 с.
Уханов В. В. Род 7. Larix Mill. – Лиственница // Деревья и кустарники СССР. Т. 1. М., Л.: Изд-во АН СССР, 1949. С. 153—176 с.
Фальк И. П. О здешних деревьях и кустах, которые годны в садах к аллеям и шпалерникам // Тр. Вольного эконом. о-ва к поощрению в России земледелия и домостроительства. Ч. 2. СПб., 1766. С. 11—32.
Фирсов Г. А., Орлова Л. В. Хвойные в Санкт-Петербурге. СПб.: ООО Издательство Росток, 2008. 336 с.
Фирсов Г. А., Фадеева И. В., Волчанская А. В. Влияние метео-фенологической аномалии зимы 2006/07 года на древесные растения в Санкт-Петербурге // Вестник МГУЛ – Лесной вестник. 2008. № 6. С. 22—27.
Фирсов Г. А., Фадеева И. В. Критические зимы в Санкт-Петербурге и их влияние на интродуцированную и местную дендрофлору // Известия Санкт-Петербургской лесотехнической академии. 2009. Вып. 188. С. 100—110.
Фирсов Г. А., Фадеева И. В., Волчанская А. В. Фенологическое состояние древесных растений в садах и парках С.-Петербурга в связи с изменениями климата // Бот. журн. 2010. Т. 95. № 1. С. 23—37.
Фирсов Г. А. Древесные растения ботанического сада Петра Великого (XVIII-XXI вв.) и климат Санкт-Петербурга // Ботаника: история, теория, практика (к 300-летию основания Ботанического института им. В. Л. Комарова Рос. акад. наук. Тр. межд. науч. конф. СПб.: Изд-во СПбГЭТУ ЛЭТИ, 2014. C. 208—215.
Фирсов Г. А., Веденяпина Е. Г., Волчанская А. В. Почвообитающие фитофторы и древесные растения в Санкт-Петербурге: новые угрозы третьего тысячелетия // Hortus Botanicus. 2014. № 9. P. 18—29. URL: http://hb.karelia.ru.
Фишер Ф. Б. Опыт разведения иностранных дерев // Лесной журнал. 1837. Ч. 3. C. 442—445.
Brasier C. M. The role of Phytophthora pathogens in forests and semi-natural communities in Europe and Africa // Phytophthora diseases of forest trees. E. M. Hansen, W. Sutton (Eds.). 2000. P. 6—13.
Dallimore W., Jackson A. B., Harrison S. G. Handbook of Conifers, including Ginkgoaceae. 4th ed. London: Edward Arnold & Co, 1966. 729 p.
Erwin D. C., Ribeiro O. K. Phytophthora diseases worldwide. St. Paul, Minnesota: APS Press, 1996. 562 p.
Grimshaw J., Bayton R. New Trees: Recent Introductions to Cultivation. The Board of Trustees of the Royal Botanic Gardens, Kew and The International Dendrology Society, 2009. 976 p.
Ho H. H., Jong S. C. Phytophthora hibernalis and P. syringae // Mycotaxon. 1993. Vol. 47. P. 439—460.
Jiménez J. J., Sánchez J. E., Romero M. A., Belbahri L., Trapero A., Lefort F., Sánchez M. E. Pathogenicity of Pythium speculum and P. sterilum on feeder roots of Quercus rotundifolia // Plant Pathology. 2008. Vol. 57. P. 369.
Jung T., Blaschke H., Oswald W. Involvment of Phytophthora species in Central European oak decline and the effect of site factors on the disease // Plant Pathology. 2000. Vol. 49. P. 706—718.
Jung T. Life cycle and pathological importance of the genus Phytophthora; URL: http://www.baumkrankheiten.com/docs-en/phytophthora.html (March 2006).
Krussmann G. Manual of Cultivated Conifers. Portland, Oregon: Timber Press, 1995. 361 p.
Manion P. D. Tree disease concepts. 2nd ed. New Jersey: Prentice-Hall. Englewood Cliffs, 1991. 402 p.
Martin F. N., Bensasson D., Tyler B. M., Boor J. L. Mitochondrial genome sequences and comparative genomics of Phytophthora ramorum and P. sojae // Curr. Genet. 2007. Vol. 51. P. 285—296.
Rehder A. A Manual of Cultivated Trees and Shrubs Hardy in North America. N., Y.: The Macmillan Company, 1949. 996 p.
Robideau G. P., De Cock A. W. A. M., Coffey M. D., Voglmayr H., Brouwer H., Bala K., Chitty D. W., Désaulniers N., Eggertson Q. A., Gachon C. M. M., Hu C.-H., Küpper F. C., Rintoul T. L., Sarhan E., Verstappen E. C. P., Zhang Y., Bonants P. J. M., Ristaino J. B., Lévesque C. A. DNAbarcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer // Molecular Ecology Resources. 2011. Vol. 11. P. 1002—1011.
Waterhouse G. M., Waterston J. M. Phytophthora cactorum // C. M. I. Descript. Pathog. Fungi Bact. 1966. Vol. 111. P. 1—2.
