

Введение
Ряд идей известного дендролога и фенолога, профессора Санкт-Петербургской государственной лесотехнической академии, Николая Евгеньевича Булыгина (1924-2002), высказанных более 40 лет назад в области интродукции растений, фенологического мониторинга и биоиндикации природных процессов особенно актуальны сейчас, на фоне глобальных изменений климата (Ярмишко, 1999; Фирсов, 2004). Уже в первых публикациях за 1961-1965 гг. Николай Евгеньевич стал интересоваться влиянием метеорологических факторов на цветение и плодоношение древесных растений, влиянием отдельных аномальных зим и лет на растения, особенностями зимнего покоя, биологии отдельных видов с целью внедрения их в озеленение Ленинграда. В конце 1960 – начале 1970-х годов вышли в свет работы об использовании математических методов в обработке материалов наблюдений, о фенопрогнозировании и закономерностях погодичного распределения фенодат (Фирсов, Ярмишко, 2005). Уже в 1970-е годы Николай Евгеньевич стал писать о потеплении климата и влиянии этого эффекта на древесные растения. В то время далеко не все исследователи всерьёз относились к этой проблеме. Влияние потепления климата на растения стало отмечаться в многочисленных научных трудах лишь годы спустя, когда проблема стала более очевидной. Выявленные Н. Е. Булыгиным климатическая и, связанная с ней, фенологическая тенденции, в последнюю четверть 20 века показали изменение уровня ритмо-адаптивных связей растений в условиях потепления климата. В результате у многих термофильных и в прошлом считавшихся непригодными для культуры в Санкт-Петербурге интродуцентов повысилась зимостойкость.
Санкт-Петербургская государственная лесотехническая академия (сейчас Санкт-Петербургский государственный лесотехнический университет имени С. М. Кирова) – известный эталонный фенологический стационар, где накоплены самые длительные в России и странах бывшего СССР ряды непрерывных фенологических наблюдений за дендрофеноиндикаторами (по зацветанию черемухи обыкновенной год начала наблюдений - 1840). Как писал сам Николай Евгеньевич, «Лесное» - старинное название северной части Санкт-Петербурга, в которой расположена Лесотехническая академия. Дендрологам сад и арборетум известен как крупный центр интродукционных испытаний древесных растений в таёжной зоне России. Фенологи считают «Лесное» старейшим в стране фенологическим стационаром. Здесь еще в 1829 г. впервые в России были начаты целенаправленные фенологические наблюдения. Несколько поколений исследователей проводят их непрерывно вот уже свыше 180 лет (Булыгин, 1980). Именно в этих непрерывных накопленных рядах и заключается отличительная особенность стационара, где работал, проводил свои наблюдения и дополнял эти ряды Н. Е. Булыгин. В некоторых странах известны и более длительные ряды наблюдений, но они охватывают всего одно-два явления и содержат много пропусков погодичных фенодат. На базе фенологического стационара «Лесное» получено 17 непрерывных рядов наблюдений длительностью свыше 75 лет по состоянию на 1995 г. (Булыгин, 1996 а), а 9 из них превышают полтора века.
Н. Е. Булыгин начал обработку этих рядов с конца 1960 гг., а первые публикации появились в 1970-х гг. Исследованиями Н. Е. Булыгина и З. Н. Довгулевич (1974 a, б), Н. Е. Булыгина (1975 а, б), Н. Е. Булыгина, С. В. Бердниковой, Л. Г. Михалкиной (1976) установлено, что в «вековых» (1841-1969 гг.) фенологических рядах, характеризующих сезонную динамику ленинградской природы от фенологического времени года «начало весны» до «апогея лета» имеется общая тенденция: наиболее позднее наступление фенодат в середине 19 века, что соответствует периоду наибольшего похолодания климата того столетия и прогрессивно раннее наступление фенодат с 90-х годов 19 века по конец 1960-х гг., как отражение известного потепления климата. Выявленную линию фенологической тенденции можно рассматривать как нисходящую ветвь достаточно длительного двухвекового фенологического цикла, являющегося зеркальным отражением восходящей ветви соответствующего климатического цикла.
Н. Е. Булыгин принял участие в Третьем совещании, посвященном ритмике природных явлений, организованном Географическим обществом СССР в 1976 г. (Булыгин, Топпер, 1976, 1977). Контрольное прогнозирование с применением фенолого-статистического метода свидетельствует о том, что при резко выраженных погодных аномалиях даже в период, для которого составлены корреляционные уравнения связи (1939-1970 гг.) фактическая ошибка прогноза хотя и не превышает средней квадратической ошибки уравнения, но всегда возрастает. Поэтому Н. Е. Булыгину представилось целесообразным проанализировать изменчивость ошибок прогноза как проявление изменчивости сопряженных фенологических лагов на 134-летних фенологических рядах за периоды, характеризующиеся наиболее поздними (1861-1880 гг.) и ранними (1921-1940 гг.) датами.
На VIII Дендрологическом конгрессе социалистических стран в Тбилиси Н. Е. Булыгин (1982 а) ввёл в научную терминологию понятие биоклиматической цикличности (БКЦ) и в дальнейшем развивал это направление науки. Под этим он понимал цикличность реакции древесных растений на короткопериодные колебания климата с учетом возрастной изменчивости самой этой реакции. Интегральным показателем БКЦ разных типов служат даты наступления фенологического периода и температуры воздуха смежных теплой и холодной частей года. Н. Е. Булыгин выделял три группы лет: ранне-тёплые, средние или нормальные и поздне-холодные. Отдельные соседние годы он объединял в циклы. Наиболее отчетливо БКЦ выражается при сопоставлении двух альтернативных биоклиматических циклов – ранне-тёплых (РТ) и поздне-холодных (ПХ), индикаторами которых служат фенодаты начала «пыления» ольхи серой (Alnus incana (L.) Moench).
Биоклиматическая цикличность – это сложное сочетание климатической и сопряженной с ней дендрофенологической цикличности в связи с многолетней изменчивостью биологических свойств растений. Именно проявление БКЦ определяет временную изменчивость как разных показателей адаптированности растений, так и оценок результатов интродукции и перспективности интродуцентов для разведения (Булыгин, 1996 б; Булыгин, Фирсов, 1998).
Н.Е. Булыгин опирался на работы своих предшественников и на достижения науки о ритмичности природных процессов. За 4 года до этого была опубликована статья Г.Е. Шульца (1978) «Фенологические наблюдения – индикатор циклических колебаний климата». Приведенные Шульцем рекогносцировочные материалы ставили на порядок дня систематические исследования по краткосрочным фенологическим, а следовательно, и климатическим циклам. К тому времени был сделан анализ на цикличность наиболее длинных многолетних рядов фенологических наблюдений. Были выявлены следующие, не остающиеся вполне постоянными циклы: 2 года, 3,5 год, 7-8 лет, 16 лет и 31 год.
Как известно, ритмичность является существеннейшим фактором изменчивости природных условий во времени и пространстве. С одной стороны, она присуща почти всем процессам в ландшафтной оболочке Земли, а с другой – именно закономерности ритмичности дают возможность предвидеть пути поступательного развития или изменчивости тех или иных интересующих нас природных явлений в перспективе (Шнитников, 1976). Явления ритмичности природных процессов привлекали и привлекают внимание всё более широких кругов учёных как в России, так и за рубежом. Как справедливо отмечает Е.В. Максимов (1976), необходимо различать три категории явлений: периодичность, цикличность и ритмичность. Первая подразумевает равновеликий характер временных интервалов. Вторая – возвращение системы в исходное положение, хотя бы и через разные промежутки времени. Третья, то есть, собственно ритмичность, напоминая одновременно и периодичность и цикличность, никогда не бывает хронологически строга и никогда не приводит систему в точности в исходное положение. В научной литературе понятия ритмичности и цикличности часто выступают как синонимы. По существу под ритмичностью подразумевают сложный и поэтому неправильный волновой процесс. Явление ритмичности относится к числу фундаментальных закономерностей природы. Ритмичность присуща широкому кругу явлений космического, геофизического и биологического характера, в том числе явлениям ландшафтной оболочки Земли в их изменчивости от нескольких лет до многих тысячелетий.
Среди внутривековых известны полтора десятка ритмов продолжительностью от 2-3 до 30-40 лет (Максимов, 1976). Установлено, что существует близкий к периодичности ритм продолжительностью примерно в 3,5 года. Однако проявляется он не на «нулевом» фоне, а интерферирует с ритмом продолжительностью в 11 лет. Именно этот последний фоновый для натурального ритм и вызывает нарушение правильности проявления натурального ритма.
Солнечная активность представляет собой сложный ритмический (полициклический) процесс, основными составляющими которого являются 5-6-летний, 11-летний, 22-летний и 80-90 летний циклы. Главный цикл – 11-летний, отчетливо видный в пятнообразовательной деятельности Солнца (числа Вольфа). Вековой, или 80-90-летний цикл модулирует амплитуды 11-летних циклов, 22-летний цикл проявляется в изменении распределения магнитных полей на Солнце, а также в чередовании более и менее интенсивных 11-летних циклов и в объединении четного и нечётного 11-летних циклов в один двойной цикл. 5-6-летний цикл виден в корпускулярной активности Солнца (возмущения магнитного поля Земли). Геомагнитные возмущения делятся на два типа: спорадические (вспышечные) и рекуррентные. Возмущения первого типа развиваются вблизи максимумов чисел Вольфа, а возмущения второго типа наиболее часты незадолго до минимумов солнечной активности. Возмущения каждого из этих типов обнаруживают 11-летнюю цикличность, но индексы магнитной возмущенности, учитывающие возмущения обоих типов, изменяются в 5-6-летнем цикле (Оль, 1976). В колебаниях атмосферной циркуляции и в обусловленных ими элементах климата (температуры, давление, осадки) отчетливо проявляются все ритмы солнечной активности, но характер этих проявлений различен в разных участках земной поверхности. Например, обнаружено, что июльские температуры в центральной части Англии в 1750-1880 гг. следовали 22-летнему циклу, причем максимальные температуры наблюдались в нечётных 11-летних циклах. Исследования колебаний атмосферных осадков и температуры при помощи спектральных методов также позволяют выделить 5-6, 11 и 22 летнюю цикличность, хотя наряду с этим отмечаются и циклы, не имеющие солнечного происхождения (Оль, 1976). Исследования других ученых показали связь засух с 11-летним циклом рекуррентных магнитных возмущений. Гелиообусловленные ритмические колебания климата приводят к соответствующим изменениям в физико-географических факторах, влияющих на состояние ландшафтной оболочки Земли. К таким факторам относятся увлажнённость земной поверхности, уровень Мирового океана и состояние ледников, уровни озер, состояние грунтовых вод, сток рек, прирост деревьев. 22-летнюю цикличность в дендрохронологических данных отмечали Т.Т. Битсвинкас, Н.В. Ловелиус, С.И. Костин, Е.В. Максимов и Н.Н. Максимова (Оль, 1976). В 1900 г. американский астроном Э.У. Браун выдвинул концепцию, согласно которой главной причиной изменения численности солнечных пятен являются приливы на Солнце, вызываемые действием приливообразующих сил Меркурия, Венеры, Земли, Юпитера и Сатурна. В частности, он показал, что учет совместного действия приливообразующих сил Юпитера и Сатурна позволяет объяснить главный период изменения численности пятен, близкий к 11 годам, и что действие сил трех других названных планет также влияет на изменение количества пятен во времени (Ржонскицкий, 1976). Выводы Брауна были подтверждены многими другими исследователями. Как отмечает С.Л. Афанасьев (1976), 11-12-летние пульсации геологических процессов на Земле хорошо увязываются с 11-летней периодичностью солнечной активности, которая, в свою очередь, по-видимому, связана с периодом обращения Юпитера вокруг Солнца (11, 86 лет). Точно так же как Луна влияет на приливные явления в атмосфере, гидросфере и литосфере Земли, планеты Солнечной системы вызывают возмущение атмосферы Солнца, усиливая процессы, приводящие к образованию солнечных пятен, и через Солнце контролирующие процессы, происходящие на Земле.
Таким образом, одним из важных результатов многолетнего фенологического мониторинга в «Лесном» явилась возможность оценки вековых изменений биоклиматических условий Петербурга и в целом всего региона под влиянием известного потепления климата в XX веке. Н.Е. Булыгиным совместно с коллегами, сотрудниками кафедры ботаники и дендрологии, преподавателями кафедры высшей математики была выполнена серия работ по математическому моделированию динамики различных фенологических процессов, феноградиентов, биофенологической, эколого-фенологической и фенолого-климатической сопряженности на основании многолетних дендрофенологических данных, накопленных в этом феностационаре и других районах России и метеорологической информации. Это позволило разработать и апробировать ряд методов долгосрочных прогнозов динамики продвижения фронта фенопроцессов, динамики наступления фенологического времени года с приуроченными к нему агротехническими мероприятиями, динамики наступления хозяйственно важных фенофаз растений. В результате стало возможным проследить современные фенологические тенденции и направления ожидаемой погоды.
Н.Е. Булыгин развил оригинальный метод фенологического прогноза теплообеспеченности вегетационного сезона, предложенный Ф.Ф. Давитая (1964). Суть его в том, что теплообеспеченность, выраженная через сумму средней суточной температуры воздуха выше 10о рассчитывается по фенодатам-индикаторам с применением соответствующего уравнения линейной регрессии. По Ф.Ф. Давитая, в районе Ленинграда выявлена поразительно высокая для биоклиматических явлений связь между весенними дендрофеноиндикаторами (начало облиствения берёзы, зацветание черёмухи и сирени) и последующей суммой температур. Как отмечает Н.Е. Булыгин с соавторами (1980, с. 62): «Климатологам ещё предстоит объяснить метеорологические причины выявленной связи». Вероятно, это связано с особенностями циркуляции атмосферы. Логично выглядит положение Ф.Ф. Давитая (1964, с. 115) о том, что на европейской территории России «как развёртывание весны, так и последующий ход погоды определяются, как правило, одними и теми же крупномасштабными процессами». Следствием этого является наличие сезонных тенденций в теплообеспеченности, о направленности которых сигнализируют растения. «С позиций интересов практики растениеводства открытого грунта (лесное и сельское хозяйство, озеленение) весьма перспективным представляется применение комбинированного прогнозирования фенологическими методами тех фенолого-метеорологических ситуаций (ФМС), в которых могут оказаться растения в тот или иной период их вегетации» (Булыгин и др., 1980, с. 65). Такое комбинированное прогнозирование заключается в сочетании фенологического расчёта динамики наступления фенофаз и ожидаемой теплообеспеченности.
Наблюдающиеся короткопериодные колебания климата Санкт-Петербурга приводят к проявлению биоклиматической цикличности (Булыгин, Бердникова, Михалкина, 1976; Фирсов, Фадеева, 2013). Цикличность обусловливает повторяемость погодичных био-метеорологических ситуаций разных типов, от благоприятных до критических по отношению к растениям. Этим в конечном итоге определяется временная амплитуда изменчивости их зимостойкости. Уровни адаптированности растений и оценки их перспективности для разведения в значительной степени связаны с тем, в биоклиматический цикл какого типа проводятся испытания (Булыгин, 1996 б; Булыгин, Фирсов, 1998). Причины противоречивости результатов исследований кроются в высокой изменчивости самой зимостойкости, одним из факторов которой является биоклиматическая цикличность. Она обуславливает повторяемость погодичных био-метеорологических ситуаций разных типов от благоприятных до критических по отношению к интродуцентам, чем в конечном итоге определяется временная амплитуда изменчивости их зимостойкости, как и других показателей уровней адаптированности. Поэтому региональный анализ проявлений биоклиматической цикличности с учетом фенологической периодизации года должен стать обязательным элементом исследований при интродукции деревянистых биоморф, особенно деревьев лесного типа и лиан. Региональный анализ проявления биоклиматической цикличности и воздействия её на интродуценты представляется необходимым элементом комплекса проводимых интродукционных испытаний, оценки перспективности интродуцентов для разведения, их репродуктивной способности, организации семеноводства и выращивания семенного потомства.
Продолжение исследований по биоклиматическому мониторингу особенно актуально в настоящее время. В условиях меняющегося климата и прогрессирующего его потепления могут значительно измениться границы агроклиматических районов и зон зимней устойчивости древесных растений. В результате чего перспективными для озеленения как Санкт-Петербурга, так и других городов России могут стать более теплолюбивые растения. Заметно может возрасти число видов и форм, пригодных для дендрологических коллекций, зимостойкость которых позволяет им расти в открытом грунте. Хотя потепление климата далеко не всегда и не во всех случаях благоприятно для интродуцированных растений, оно открывает более широкие возможности как для первичной, так и повторной интродукции. Это дает возможность выращивать больше видов из семян местной репродукции и способствовать в конечном счете их акклиматизации.
Основанием для отнесения каждого года к тому или иному биоклиматическому циклу служат статистики ряда распределения фенодат зацветания Alnus incana. В практике интродукционных испытаний, проводимых в С.-Петербурге, в последние десятилетия показатели адаптированности древесных интродуцентов оцениваются через показатели биоклиматической цикличности по трём обусловленным ею уровням биоклиматической ситуации: типичной для подзоны южной тайги; благоприятной (с повышенной теплообеспеченностью холодных и теплых сезонов года) и неблагоприятной. Наиболее отчетливо биоклиматическая изменчивость выражается при сопоставлении двух альтернативных биоклиматических циклов – ранне-теплого и поздне-холодного. Проявление цикличности (которая существовала, очевидно, всегда, при любом климате), сейчас происходит на фоне климатической тенденции в направлении его потепления.
Для нас представляет интерес, как проявляется биоклиматическая цикличность сейчас, в начале третьего тысячелетия, насколько различаются метеорологические показатели в ранне-теплые и поздне-холодные годы. Какова реакция растений, проявляется ли она в годы с разной биоклиматической ситуацией и насколько. Этому и посвящена наша статья.
Принятые обозначения: ПВ – подсезон «Предвесенье», СТ1 – первый феноэтап подсезона «Снеготаяние», СТ2 – второй феноэтап подсезона «Снеготаяние», ОВ1 и ОВ2 – первый и второй феноэтапы подсезона «Оживление весны», РВ1 и РВ2 – первый и второй феноэтапы подсезона «Разгар весны», НЛ1 –первый феноэтап подсезона «Начало лета», НО1 –первый феноэтап подсезона «Начало осени», ЗО1 и ЗО2–первый и второй феноэтапы подсезона «Золотая осень», ГО1 и ГО2 – первый и второй феноэтапы подсезона «Глубокая осень», ПЗ – подсезон «Предзимье», ПРз – «Первозимье» (начало зимы).
Объекты и методы исследований
Объектами исследования являются древесные растения интродуцированной и местной дендрофлоры в Санкт-Петербурге и дендрофеноиндикаторы Календаря природы. Исследование выполнено в феностационаре Ботанического сада Петра Великого БИН РАН. Использованы литературные данные по интродукции древесных растений и результаты собственных наблюдений. Ежегодная оценка обмерзания проводится с начала 1980-х гг. по шкале П. И. Лапина (1967): 1 — отсутствие повреждений, 2 — подмерзание хвои и концов однолетних побегов, 7 — гибель растения от морозов. В работе использованы данные метеостанции Санкт-Петербург Северо-Западного межрегионального территориального управления федеральной службы по гидрометеорологии и мониторингу окружающей среды. Обозначения фенофаз даны по Н. Е. Булыгину (1979). Естественная периодизация года приводится по Н. Е. Булыгину (1982 а). Деление на ранне-теплые и поздне-холодные годы выполнено по методике Н. Е. Булыгина (1982 б).
Результаты и обсуждение
По методике Н. Е. Булыгина были выделены ранне-тёплые и поздне-холодные годы за период 1980-2021 гг. (42 года). Средняя дата зацветания ольхи серой за период 1980-2021 гг. (n=42) - 1 апреля ± 2,1 сут. Если принять значение ±3 m за норму (при P=0,99), то в норму попадают годы: 1 апреля ± 6,5 сут., то есть, от 26 марта по 8 апреля включительно. Годы с зацветанием ольхи 25 марта и ранее, попадают в «ранние» и, соответственно, от 9 апреля и позже - в поздние (рис. 1). Оказалось, что к поздне-холодным годам можно отнести 12 лет: 1980, 1982, 1985, 1987, 1996, 1998, 2003, 2004, 2006, 2011, 2012 и 2013 гг. А к ранне-тёплым: 1989, 1990, 1992, 1995, 2007, 2008, 2014, 2015, 2016, 2019 и 2020 гг. (11 лет).
На рис. 1 обозначено: Х - среднее значение ряда дат зацветания Alnus incana.
Различие в температурном режиме по месяцам и годам в ранне-тёплые и поздне-холодные годы можно видеть из данных таблицы 1.
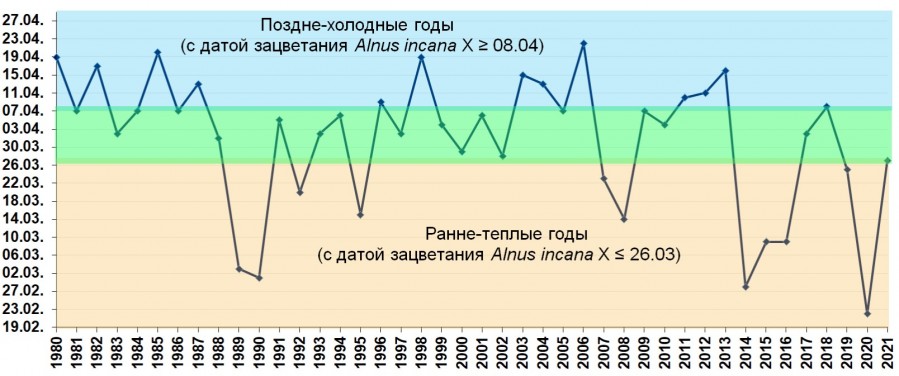
Рис. 1. Даты зацветания Alnus incana (1830-2021 гг.) – индикатор короткопериодных колебаний климата.
Таблица 1. Среднемесячные и годовая температура воздуха в Санкт-Петербурге в ранне-тёплые и поздне-холодные годы (1980-2021 гг.).
| Годы | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | Год |
| ПОЗДНЕ-ХОЛОДНЫЕ ГОДЫ (n=12) | |||||||||||||
| 1980 | -8,9 | -8,3 | -4,6 | 5,0 | 7,7 | 18,6 | 17,0 | 15,5 | 11,1 | 5,9 | -1,7 | -3,4 | 4,5 |
| 1982 | -10,6 | -6,8 | -0,1 | 4,1 | 10,4 | 11,9 | 17,7 | 16,6 | 11,2 | 5,3 | 3,7 | -0,3 | 5,3 |
| 1985 | -13,7 | -14,9 | -0,8 | 2,3 | 10,1 | 13,4 | 16,4 | 18,4 | 10,9 | 7,5 | -1,0 | -6,0 | 3,6 |
| 1987 | -17,9 | -5,6 | -4,7 | 2,6 | 10,2 | 14,9 | 15,9 | 13,9 | 9,8 | 6,4 | -2,0 | -5,5 | 3,2 |
| 1996 | -7,0 | -10,9 | -2,9 | 3,4 | 10,3 | 14,7 | 16,2 | 18,4 | 9,7 | 6,6 | 4,0 | -5,2 | 4,8 |
| 1998 | -1,7 | -7,3 | -4,2 | 3,6 | 10,8 | 16,4 | 18,0 | 14,7 | 11,8 | 6,5 | -5,1 | -4,1 | 5,0 |
| 2003 | -9,5 | -5,7 | -1,6 | 2,3 | 12,1 | 13,0 | 21,4 | 16,6 | 12,4 | 5,1 | 2,4 | -0,8 | 5,6 |
| 2004 | -8,1 | -5,4 | -0,6 | 4,9 | 11,1 | 14,7 | 18,5 | 17,8 | 13,2 | 6,5 | -0,1 | -0,7 | 6,0 |
| 2006 | -5,8 | -10,7 | -5,8 | 5,4 | 11,8 | 17,0 | 19,2 | 18,9 | 14,2 | 8,1 | 1,7 | 3,0 | 6,4 |
| 2011 | -5,8 | -11,0 | -1,9 | 5,7 | 11,0 | 17,7 | 22,5 | 17,5 | 13,1 | 7,6 | 3,6 | 1,9 | 6,8 |
| 2012 | -4,8 | -10,4 | -1,0 | 4,9 | 12,7 | 15,2 | 19,5 | 16,3 | 12,9 | 6,6 | 2,9 | -7,9 | 5,6 |
| 2013 | -6,1 | -2,6 | -6,6 | 4,2 | 14,4 | 19,8 | 19,0 | 18,6 | 12,1 | 7,3 | 4,4 | 0,9 | 7,1 |
| X | -8,3 | -8,3 | -2,9 | 4,0 | 11,1 | 15,6 | 18,4 | 16,9 | 11,9 | 6,6 | 1,1 | -2,3 | 5,2 |
| РАННЕ-ТЁПЛЫЕ ГОДЫ (n=11) | |||||||||||||
| 1989 | -0,7 | 0,6 | 2,5 | 7,7 | 12,5 | 18,4 | 18,9 | 16,4 | 12,9 | 6,0 | 0,6 | -5,0 | 7,6 |
| 1990 | -5,4 | 1,7 | 1,4 | 8,1 | 9,8 | 14,3 | 17,0 | 16,6 | 9,0 | 5,8 | -0,5 | -1,8 | 6,3 |
| 1992 | -2,7 | -2,7 | 1,8 | 3,3 | 11,0 | 16,7 | 17,8 | 16,6 | 13,8 | 1,5 | -2,5 | -0,1 | 6,2 |
| 1995 | -4,1 | -0,3 | 1,1 | 4,8 | 11,1 | 19,3 | 16,5 | 16,9 | 12,3 | 8,3 | -1,8 | -8,6 | 6,3 |
| 2007 | -2,8 | -10,6 | 3,6 | 5,2 | 12,4 | 16,0 | 18,5 | 19,6 | 12,1 | 7,5 | -0,8 | 0,9 | 6,8 |
| 2008 | -1,8 | -0,2 | 0,4 | 7,0 | 11,0 | 15,1 | 17,9 | 16,0 | 10,7 | 8,8 | 3,0 | -0,9 | 7,3 |
| 2014 | -7,0 | 0,0 | 2,2 | 6,5 | 13,0 | 15,0 | 21,2 | 18,8 | 13,5 | 5,2 | 0,8 | -1,0 | 7,4 |
| 2015 | -2,7 | -0,6 | 2,6 | 5,1 | 11,8 | 15,9 | 16,9 | 18,2 | 14,0 | 5,6 | 3,1 | 2,1 | 7,7 |
| 2016 | -11,2 | 0,0 | 1,0 | 6,3 | 14,7 | 16,4 | 19,0 | 17,2 | 12,9 | 5,0 | -1,8 | -1,2 | 6,5 |
| 2019 | -6,4 | -0,5 | 0,1 | 7,3 | 12,1 | 18,6 | 16,6 | 17,0 | 12,2 | 6,1 | 1,9 | 1,8 | 7,2 |
| 2020 | 1,5 | 0,6 | 2,2 | 4,2 | 10,0 | 19,1 | 17,6 | 17,2 | 14,3 | 9,1 | 3,9 | -0,7 | 8,3 |
| Х | -3,9 | -1,1 | 1,7 | 5,9 | 11,8 | 16,8 | 18,0 | 17,3 | 12,5 | 6,3 | 0,5 | -1,3 | 7,1 |
| ∆Х | +4,4 | +7,2 | +4,6 | +1,9 | +0,7 | +1,2 | -0,4 | +0,4 | +0,6 | -0,3 | -0,6 | +1,0 | 1,9 |
| Годы | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | Год |
Примечание: В таблице 1 обозначено: Х - среднее значение, ∆Х - разница температур между ранне-тёплыми и поздне-холодными годами.
Таблица 2. Даты последнего весеннего и первого осеннего заморозков в воздухе в поздне-холодные и ранне-теплые годы, 1980-2021 гг.
| Год | Последний весенний заморозок в воздухе | Первый осенний заморозок в воздухе | Безмо-розный период, сут. | ||||
| дата | феноэтап | То, С | дата | феноэтап | То, С | ||
| ПОЗДНЕ-ХОЛОДНЫЕ ГОДЫ, n=12 | |||||||
| 1980 | 22 мая | РВ1 | -1,2 | 24 октября | ПЗ | -2,2 | 155 |
| 1982 | 23 апреля | ОВ1 | -0,4 | 11 октября | ЗО2 | -2,4 | 171 |
| 1985 | 22 мая | РВ1 | -0,3 | 18 октября | ЗО2 | -2,1 | 149 |
| 1987 | 28 апреля | ОВ2 | -0,3 | 21 октября | ГО2 | -0,3 | 176 |
| 1996 | 25 апреля | ОВ1 | -2,8 | 15 октября | ЗО2 | -1,0 | 173 |
| 1998 | 16 апреля | СТ2 | -3,1 | 06 октября | ЗО1 | -1,6 | 173 |
| 2003 | 27 апреля | ОВ2 | -3,8 | 19 октября | ГО1 | -2,6 | 175 |
| 2004 | 13 мая | РВ2 | -0,6 | 10 октября | ЗО2 | -0,2 | 150 |
| 2006 | 24 апреля | ОВ1 | -0,8 | 29 октября | ГО1 | -1,8 | 188 |
| 2011 | 20 апреля | ОВ1 | -0,7 | 15 октября | ЗО2 | -0,8 | 178 |
| 2012 | 19 апреля | ОВ1 | -1,2 | 26 октября | ГО1 | -1,2 | 190 |
| 2013 | 29 апреля | ОВ2 | -0,5 | 14 октября | ГО1 | -0,2 | 168 |
| Х | 30 апреля | - | - | 17 октября | - | - | 170 |
| РАННЕ-ТЕПЛЫЕ ГОДЫ, n=11 | |||||||
| 1989 | 06 апреля | ОВ1 | -1,9 | 19 октября | ГО1 | -1,0 | 196 |
| 1990 | 12 мая | РВ2 | -0,2 | 03 октября | ЗО2 | -1,7 | 144 |
| 1992 | 25 апреля | ОВ2 | -1,0 | 11 октября | ПЗ | -1,5 | 169 |
| 1995 | 11 мая | РВ1 | -0,9 | 22 октября | ГО2 | -1,5 | 164 |
| 2007 | 01 мая | РВ1 | -1,3 | 04 ноября | ПЗ | -1,5 | 187 |
| 2008 | 20 апреля | ОВ2 | -0,5 | 07 ноября | ПЗ | -1,5 | 201 |
| 2014 | 10 апреля | ОВ1 | -2,4 | 17 октября | ЗО2 | -1,6 | 190 |
| 2015 | 19 апреля | ОВ2 | -1,2 | 07 октября | ЗО1 | -0,2 | 171 |
| 2016 | 12 апреля | ОВ2 | -0,5 | 19 октября | ГО1 | -0,4 | 190 |
| 2019 | 14 апреля | ОВ2 | -1,3 | 06 октября | ЗО2 | -2,0 | 175 |
| 2020 | 25 апреля | ОВ2 | -1,0 | 21 октября | ЗО2 | -0,9 | 179 |
| Х | 22 апреля | - | - | 18 октября | - | - | 179 суток |
Примечание: В таблице 2 обозначено: Х - среднее значение.
Среднегодовая температура в ранне-тёплые годы на 1,9о больше, чем в поздне-холодные (увеличилась с 5,2о до 7,1о). Это очень сильное различие и высокие абсолютные значения температуры. Можно вспомнить, что во второй половине 20 века «нормой современного климата» в Ленинграде считалась годовая температура 4,3о. Если посмотреть по месяцам, то очень сильно различаются два самых холодных месяца года: январь - на 4,4о и февраль - на 7,2о. Март в ранне-тёплые годы во всех случаях стал уже не зимним, а с положительной температурой воздуха, и отличается от поздне-холодных лет на 4,6о. Достаточно высокое различие и в апреле (на 1,9о). Оно проявляется по июнь включительно (1,2о), в другие месяцы различие уже незначительное. Из данных таблицы 1 видно важное значение режима температуры для растений в ранне-тёплые годы по сравнению с поздне-холодными. В ранне-тёплые годы более длинный вегетационный сезон, лучшие условия для перезимовки древесных растений, вызревания их побегов и подготовки к зиме. Важно обратить внимание, что более высокая обеспеченность теплом в ранне-тёплые годы проявляется в самые холодные зимние месяцы года и в первой половине вегетационного сезона.
Можно заметить, что после тёплых зим, какими были зимы 1988/89 и 1989/90 гг. у 70-80 % видов коллекции Ботанического сада Петра Великого обмерзания отсутствовали, у 10-15 % были отмечены повреждения почек и концов побегов. И лишь у отдельных видов имело место обмерзание побегов старше одного года (Фирсов и др., 2008).
В таблице 2 приводятся даты последних весенних и первых осенних заморозков, а также продолжительность безморозного сезона в поздне-холодные и ранне-тёплые годы.
Оказывается, что различие существует, и в ранне-тёплые годы весенние заморозки оканчиваются на 8 суток раньше, а в целом продолжительность безморозного периода увеличивается на 9 суток, что является благоприятным фактором для растений.
Если мы сравним динамику сезонного развития по Календарю природы в ранне-тёплые и поздне-холодные годы, то увидим значительную разницу между этими группами лет. В среднем в ранне-тёплые годы от весны и до самой осени даты наступления феноэтапов года идут с опережением. В конце зимы и начале весны различие очень сильное, но оно не исчезает и до осени. Различие на этапе «Предвесенье» - 41 сут., на первом этапе подсезона «Снеготаяние» - 37 сут., а на втором его этапе - 26 сут., на первом этапе подсезона «Оживление весны» - 36 сут. То есть, ольха серая зацветает в ранне-тёплые годы в среднем раньше, чем в поздне-холодные годы более чем на месяц. На втором этапе подсезона «Оживления весны» различие несколько сглаживается - до 15 сут. И далее, от подсезона «Разгар весны» и до подсезона «Начало осени», различие хотя и не так сильно выражено, но сохраняется: от 5 суток на третьем этапе подсезона «Разгар весны» до 15 суток на первом этапе подсезона «Спад лета». В поздне-холодные годы сезонное развитие природы идёт, как правило, с отставанием от среднемноголетних сроков. В 1980 г. 14 этапов до самого конца лета были достоверно поздними, и лишь один (второй этап подсезона «Начало лета») попал в средние сроки. Такое же выраженное холодное лето с поздними датами от начала подсезона «Оживление весны» до второго этапа подсезона «Спад лета» включительно (12 феноэтапов подряд) было в 1982 г. В 1985 г. с поздними датами наступления было 13 этапов подряд до самого начала осени. В холодный 1987 г., когда в Ленинградской области местами ушла в зиму невызревшая клюква (Фирсов, Фадеева, 2009), тоже имели место отрицательные аномалии: 7 феноэтапов подряд, от второго этапа подсезона «Разгар весны» до третьего этапа подсезона «Полное лето», а также второй этап подсезона «Спад лета». В годы ранне-тёплые, наоборот, развитие природы в подавляющем большинстве случаев идёт с опережением своих среднемноголетних значений. Очень выражен при этом год 1989, с которого началось заметное потепление климата в Санкт-Петербурге (Фирсов, 2014; Фирсов, Волчанская, 2021) - все феноэтапы года от «Предвесенья» и до начала осени были ранними.
Представляет интерес проследить сроки наступления и продолжительность сезонов года в ранне-тёплые и поздне-холодные годы (табл. 3).
Таблица 3. Даты наступления и продолжительность сезонов года в Ботаническом саду Петра Великого в Санкт-Петербурге, годы РТ и ПХ (1980-2021 гг.)
| Годы | Перво-зимье (ПРз) пред-шествую-щей зимы | Снего-таяние первый этап (СТ1) | Дли-тель-ность зимы | Начало лета первый этап (НЛ1) | Дли-тель-ность вес-ны | Начало осени первый этап (НО1) | Дли-тель-ность лета | Перво-зимье (ПРз) | Дли-тель-ность осени |
| РАННЕ-ТЕПЛЫЕ ГОДЫ, n=11 | |||||||||
| 1989 | 30 октября | 26 января | 88 | 24 мая | 119 | 23 августа | 91 | 16 ноября | 85 |
| 1990 | 16 ноября | 29 января | 74 | 26 мая | 118 | 26 августа | 92 | 12 ноября | 78 |
| 1992 | 05 декабря | 29 февраля | 86 | 08 июня | 100 | 30 августа | 83 | 24 октября | 55 |
| 1995 | 03 ноября | 14 февраля | 103 | 31 мая | 107 | 20 августа | 81 | 01 ноября | 73 |
| 2007 | 20 января | 02 марта | 41 | 31 мая | 90 | 05 сентября | 97 | 05 ноября | 61 |
| 2008 | 05 ноября | 07 марта | 123 | 30 мая | 84 | 02 сентября | 95 | 08 декабря | 97 |
| 2014 | 11 января | 09 февраля | 29 | 25 мая | 106 | 02 сентября | 100 | 22 декабря | 111 |
| 2015 | 22 декабря | 20 февраля | 60 | 04 июня | 105 | 27 августа | 84 | 27 декабря | 122 |
| 2016 | 27 декабря | 27 января | 31 | 25 мая | 119 | 27 августа | 94 | 02 ноября | 67 |
| 2019 | 26 ноября | 15 марта | 109 | 27 мая | 73 | 25 августа | 90 | 26 января | 154 |
| 2020 | 26 января | 09 февраля | 14 | 4 июня | 116 | 01 сентября | 89 | 07 декабря | 97 |
| Х | 09 декабря | 16 февраля | 69 | 30 мая | 103 | 28 августа | 91 | 27 ноября | 91 |
| ПОЗДНЕ-ХОЛОДНЫЕ ГОДЫ, n=12 | |||||||||
| 1980 | 29 ноября | 28 марта | 120 | 10 июня | 74 | 24 августа | 75 | 29 октября | 66 |
| 1982 | 12 ноября | 13 марта | 121 | 15 июня | 94 | 26 августа | 72 | 08 декабря | 104 |
| 1985 | 10 ноября | 14 марта | 124 | 15 июня | 93 | 04 сентября | 81 | 16 ноября | 73 |
| 1987 | 03 декабря | 22 марта | 109 | 11 июня | 81 | 30 августа | 80 | 07 ноября | 69 |
| 1996 | 01 ноября | 24 марта | 144 | 09 июня | 77 | 08 сентября | 91 | 11 декабря | 94 |
| 1998 | 17 ноября | 27 марта | 130 | 08 июня | 73 | 03 сентября | 87 | 08 ноября | 66 |
| 2003 | 03 ноября | 23 марта | 140 | 09 июня | 78 | 03 сентября | 86 | 06 декабря | 94 |
| 2004 | 06 декабря | 13 марта | 98 | 05 июня | 84 | 30 августа | 86 | 17 ноября | 79 |
| 2006 | 04 декабря | 29 марта | 115 | 04 июня | 67 | 26 августа | 83 | 20 января | 147 |
| 2011 | 21 ноября | 01 апреля | 131 | 03 июня | 63 | 26 августа | 84 | 01 января | 128 |
| 2012 | 01 января | 02 апреля | 92 | 01 июня | 60 | 28 августа | 88 | 28 ноября | 92 |
| 2013 | 28 ноября | 04 апреля | 127 | 31 мая | 57 | 01 сентября | 93 | 11 января | 132 |
| X | 24 ноября | 24 марта | 120 | 08 июня | 75 | 30 августа | 84 | 04 декабря | 95 |
В таблице 3 обозначено: Х - среднее значение.
Благоприятным для растений является более продолжительный период вегетации, наблюдающийся в ранне-тёплые циклы. Удлинение вегетационного сезона происходит в первую очередь за счет более раннего его начала в ранне-теплые годы, что обусловлено самим принципом их выделения, основанным на отклонении в сроках зацветания ольхи серой от нормы. Кроме того, в ранне-теплые годы на неделю позже наступает феноэтап «Первозимье». Обращает на себя внимание, что в ранне-тёплые годы резко сокращается продолжительность зимы, в среднем на 52 сут. или почти на два месяца. Сюда попали все очень короткие и рекордные годы, когда продолжительность зимнего периода была минимальной (зима 2019-2020 гг. - 14 сут.). Наоборот, в поздне-холодные годы зимний период удлиняется (до 140 сут. зимой 2002/03). В годы ранне-тёплые заметно удлиняется весенний период, в среднем на 28 сут. и увеличивается продолжительность лета - за счет более раннего его начала, в среднем на 7 сут. Заметно отличаются даты наступления сезонов года. Так, в ранне-теплые годы весна начинается в среднем 16 февраля, а в годы поздне-холодные - 24 марта. Осенние периоды близки между собой (продолжительность 91 и 95 сут. соответственно). Можно заметить, что на динамику осенних фенофаз влияют другие закономерности. Ритмика отмирания ассимиляционного аппарата осенью больше подвержена воздействию заморозков, а не общего хода снижения температуры (Фирсов, Фадеева, 2013). В целом осенью коррелирует с биоритмикой весенне-летних подсезонов только динамика созревания плодов. В ритмике отмирания листьев нет таких корреляций с предшествующими феноэтапами года, на что обращал внимание ещё Н. Е. Булыгин (1980).
Начиная с XIX века много попыток было сделано, чтобы распределить растения на категории устойчивости для садоводческих целей. Проблема районирования территории России с точки зрения ассортимента интродуцированных древесных растений, устойчивых в той или иной зоне, не нова и ей уделялось внимание дендрологами России и бывшего СССР. В связи с интенсивной интродукцией инорайонного растительного материала вопрос об устойчивости и классификации интродуцентов по климатическим зонам широко обсуждался и обсуждается и среди зарубежных дендрологов (Magarey et al., 2008; Prokhorov et al., 2012; Daly et al., 2012). Карты зон устойчивости основываются на климатических параметрах, обычно по зимним изотермам (линиям с одинаковыми температурами воздуха). Соответственно, области со сходным климатом объединяются вместе. Они неизбежно слишком широкие и приблизительные. Микроклимат в этом случае вообще не учитывается. Но в целом такая классификация – полезное руководство для общих представлений и предварительного отбора растений. Величины, близкие к абсолютному минимуму, встречаются редко, один раз в 50-80 лет, поэтому на практике агроклиматологии в качестве показателя морозоопасности пользуются средними из абсолютных минимумов температуры воздуха. Этот же климатический показатель был принят за основу Альфредом Редером (Rehder, 1949), чей широко известный справочник до сих пор является настольной книгой для дендрологов стран умеренного климата. Справочными данными Редера до сих пор пользуются российские дендрологи для предварительной оценки адаптационных возможностей интродуцентов, так как Редер испытал наибольшее число древесных видов в мире на тот момент, и его данные очень полные и до сих пор актуальные. В его справочнике приводится карта территории США и Канады с выделением 7 зон зимней устойчивости древесных растений. Для более чем 2550 видов деревьев и кустарников отмечается возможная зона их культуры в открытом грунте. Зоны выделены Департаментом сельского хозяйства США, основываясь на средней из минимальных температур за период 20 лет. Позже эта система была повторно проанализирована, уточнена и дополнена (Warda, 1995; Beckett, 1995). Сейчас признаются 11 зон: зона 1 – Арктика, 10 и 11 – тропики (южная часть п-ова Флорида). В последние десятилетия система зон устойчивости была распространена и на Западную Европу. Садоводы и дендрологи в США и Канаде вскоре после ее создания приняли концепцию зон устойчивости растений со всеми ее преимуществами и недостатками. При этом нельзя недооценивать влияние на растения и многих других факторов, как-то: летнее тепло, годовое количество осадков и их распределение по сезонам, выпадение зимних осадков в виде снега, интенсивность зимнего солнца, ветер и различные почвенные факторы. Некоторые интродуценты, как например, виды семейства Ericaceae, требовательны к кислотности почвы, в противном случае они могут погибнуть, невзирая на самый подходящий климат. При культуре низкорослых растений на садовых участках, особо ценных экземпляров в арборетумах и ботанических садах, возможно повышение их устойчивости за счет укрытия, выбора мест посадки, надлежащей агротехники. Во всех же остальных случаях нужно ориентироваться на проверенные и зимостойкие растения (Фирсов, 2003; Фирсов, Фадеева, 2020). Таким образом, самая широко используемая карта зон устойчивости – это та, что разработана Департаментом сельского хозяйства США (US Department of Agriculture, USDA). Она основывается на очень простой идее, используя среднеминимальные температуры воздуха, разделённые на изотермы через 10о по шкале Фаренгейта. Иногда подразделяется на интервалы через 5 оF по той же шкале, с наименованием «а» для более холодной половины зоны и «b» - для более тёплой подзоны. Европейцы уже давно перевели градусы по шкале Фаренгейта в более понятную для себя шкалу Цельсия (таблица 4).
Таблица 4. Зоны устойчивости древесных растений Департамента сельского хозяйства США
| Зона устойчивости | То по шкале Фаренгейта | То по шкале Цельсия |
| 1 | Ниже -50 o F | Ниже -45,6 oC |
| 2a | -50 до -45 оF | -45,5 до -42,8 oC |
| 2b | -45 до -40 оF | -42,7 до -40,0 oC |
| 3a | -40 до -35 оF | -39,9 до -37,3 oC |
| 3b | -35 до -30 оF | -37,2 до -34,5 oC |
| 4а | -30 до -25 оF | -34,4 до -31,7 oC |
| 4b | -25 до -20 оF | -31,6 до -28,9 oC |
| 5а | -20 до -15 оF | -28,8 до -26,2 oC |
| 5b | -15 до -10 оF | -26,1 до -23,4 oC |
| 6а | -10 до -5 оF | -23,3 до -20,6 oC |
| 6b | -5 до 0 оF | -20,5 до -17,8 oC |
| 7а | 0 до 5 оF | -17,7 до -15,0 oC |
| 7b | 5 до 10 оF | -14,9 до -12,3 oC |
| 8а | 10 до 15 оF | -12,2 до -9,5 oC |
| 8b | 15 до 20 оF | -9,4 до -6,7 oC |
| 9а | 20 до 25 оF | -6,6 до -3,9 oC |
| 9b | 25 до 30 оF | -3,8 до -1,2 oC |
| 10а | 30 до 35 оF | -1,1 до 1,6 oC |
| 10b | 35 до 40 оF | 1,7 до 4,4 oC |
| 11 | Выше 40 оF | Выше 4,5 oC |
Эту систему уже давно применяют в Австралии и Новой Зеландии. И за многие годы, во многом путем прямых проб и ошибок, большинство видов деревьев и кустарников были оценены с точки зрения их отнесения к той или иной зоне (Фирсов, Фадеева, 2020). Известно, что губительное действие на растения оказывает не средний уровень зимних температур, а наиболее суровые, хотя и кратковременные морозы. В таблице 5 приводится абсолютно минимальная температура воздуха в Санкт-Петербурге за каждый из ранне-тёплых и поздне-холодных лет.
В нашем случае разница в среднемноголетнем значении в 7,2о очень заметная. Это означает, что С.-Петербург в ранне-тёплые годы находится в зоне 6, в более тёплой её подзоне 6 б, а в поздне-холодные годы перемещается в зону 5, в более холодную подзону 5а совсем с другими условиями перезимовки и существования растений. Это, конечно, может сказаться и на состоянии растений, и на результатах их перезимовки в разные биоклиматические циклы. В этих зонах предполагается и разный ассортимент деревьев и кустарников для разведения. В случае дальнейшего потепления климата при преодолении рубежа 17,8о (до чего осталось 1,5о), С.-Петербург перейдёт в 7 зону, куда входят многие виды деревьев и кустарников, которые сейчас считаются оранжерейно-комнатными растениями. И целый ряд из них даже и не пытались испытывать в открытом грунте. Можно заметить, что минимальная температура -8,9о зимой 2019/20 г. - абсолютный рекорд за весь период наблюдений. В эту зиму в Ботаническом саду Петра Великого перезимовал даже Callitris oblonga A. et L. C. Rich., представляющий флору Австралии (вымерз следующей зимой).
В таблицах 6 и 7 приводятся списки видов, вымерзших в парке-дендрарии БИН РАН в годы поздне-холодные и ранне-тёплые в 21 веке по актам списания. Интересно посмотреть, есть ли в этом случае различие, и в какой степени оно проявляется.
Таблица 5. Абсолютный минимум температуры воздуха в Санкт-Петербурге в ранне-тёплые и поздне-холодные годы в период 1980-2021 гг.
| РАННЕ-ТЁПЛЫЕ ГОДЫ, n=11 | ПОЗДНЕ-ХОЛОДНЫЕ ГОДЫ, n=12 | ||||||
| Год | Зимний период | Т оС | Дата | Год | Зимний период | Т оС | Дата |
| 1989 | 1988-89 | -23,1 | 02.01.89 | 1980 | 1979-80 | -25,8 | 11.02.80 |
| 1990 | 1989-90 | -22,9 | 15.01.90 | 1982 | 1981-82 | -27,6 | 10.01.82 |
| 1992 | 1991-92 | -18,1 | 21.02.92 | 1985 | 1984-85 | -30,0 | 21.01.85 |
| 1995 | 1994-95 | -19,1 | 19.12.94 | 1987 | 1986-87 | -34,7 | 10.01.87 |
| 2007 | 2006-07 | -22,8 | 08.02.07 | 1996 | 1995-96 | -26,7 | 10.02.96 |
| 2008 | 2007-08 | -15,2 | 06.01.08 | 1998 | 1997-98 | -22,9 | 15.12.97 |
| 2014 | 2013-14 | -20,3 | 23.01.14 | 2003 | 2002-03 | -28,8 | 11.01.03 |
| 2015 | 2014-15 | -14,7 | 22.01.15 | 2004 | 2003-04 | -21,5 | 21.01.04 |
| 2016 | 2015-16 | -24,5 | 08.01.16 | 2006 | 2005-06 | -27,8 | 19.01.06 |
| 2019 | 2018-19 | -22,5 | 22.01.19 | 2011 | 2010-11 | -26,8 | 18.02.11 |
| 2020 | 2019-20 | -8,9 | 05.02.20 | 2012 | 2011-12 | -22,5 | 02.02.12 |
| - | - | - | - | 2013 | 2012-13 | -22,3 | 19.01.13 |
| Х | - | -19,3 | - | Х | - | -26,5 | - |
В таблице 5 обозначено: Х - среднее значение.
Таблица 6. Древесные растения, погибшие в Ботаническом саду Петра Великого в поздне-холодные годы 21 века
| Название растений | Участок | 2003 | 2004 | 2006 | 2011 | 2012 | 2013 |
| Acanthopanax sessiliflorus (Rupr. et Maxim.) Seem. | 18 | - | - | - | + | - | - |
| Acanthopanax simonii (Decne.) C. K. Schneid. | 91 | - | -- | - | - | - | + |
| Acer cissifolium (Siebold et Zucc.) C. Koch | 9 | - | -- | - | + | - | - |
| Acer glabrum Torr. | 83 | - | - | + | - | - | - |
| Acer saccharum Marshall subsp. nigrum (Michx. f.) Desmarais | 1 | + | - | - | - | - | - |
| Acer sieboldianum Miq. | 19 | - | - | + | - | - | - |
| Acer triflorum Kom. | 19 | - | - | + | - | - | - |
| Acer velutinum Boiss. | 19 | - | + | - | - | - | - |
| Artemisia santonica L. | 99 | - | - | - | - | + | - |
| Atragene speciosa Weinm. | 101 | - | - | - | - | + | - |
| Berberis sphaerocarpa Kar. et Kir. | 49 | - | - | + | - | - | - |
| Berberis x emarginata Willd. | 37 | - | - | - | + | - | - |
| Caragana pygmaea (L.) DC. | 7 | - | - | - | + | - | - |
| Chamaedaphne calyculata (L.) Moench | 99 | - | - | - | - | + | - |
| Cotoneaster multiflorus Bunge | 26 | - | + | - | - | - | - |
| Cotoneaster roseus Edgew. | 11 | - | - | + | - | - | - |
| Crataegus champlainensis Sarg. | 42 | - | + | - | - | - | - |
| Crataegus remotilobata Raikova ex Popov | 8 | - | - | - | - | + | - |
| Deutzia x rosea (Lemoine) Rehd. | 17 | - | + | - | - | - | - |
| Deutzia scabra Thunb. | 67 | - | + | - | - | - | - |
| Erica scoparia L. | 99 | - | - | - | - | - | + |
| Exochorda korolkowii Lavall. | 101 | - | - | + | - | - | - |
| Exochorda giraldii Hesse | 51 | - | - | - | - | - | + |
| Fagus orientalis Lipsky | 17 | - | - | + | - | - | - |
| Fraxinus quadrangulata Michx. | 12 | - | + | - | - | - | - |
| Laburnum anagyroides Medik. ‘Alschingeri’ | 13 | - | + | - | - | - | - |
| Laburnum anagyroides Medik. ‘Quercifolia’ | 50 | + | - | - | - | - | - |
| Lespedeza bicolor Turcz. | 91 | - | - | - | - | + | - |
| Ligusrum ovalifolium Hassk. | 1 | + | - | - | - | - | - |
| Ligustrum ovalifolium Hassk. | 17 | - | + | - | - | - | - |
| Lonicera dioica L. | 10 | - | - | - | + | - | - |
| Lonicera longipes (Maxim.) Pojark. | 94 | - | + | - | - | - | - |
| Osmaronia cerasiformis (Torr. et Gray) Greene | 17 | - | - | + | - | - | - |
| Persica vulgaris Mill. ‘Plena’ | 94 | - | - | - | - | + | |
| Philadelphus purpurascens (Koehne) Rehder | 50 | - | - | + | - | - | - |
| Pinus armandii Franch. | 98 | + | - | - | - | - | - |
| Platycladus orientalis (L.) Franco | 142 | + | - | - | - | - | - |
| Platycladus orientalis (L.) Franco | 91 | + | - | - | - | - | - |
| Populus tremula L. var. davidiana (Dode) C. K. Schneid. | 22 | - | - | - | - | + | - |
| Prunus sogdiana Vass. | 13 | - | - | + | - | - | - |
| Prunus sogdiana Vass. | 8 | - | - | + | - | - | - |
| Pterostyrax hispida Siebold et Zucc. | 91 | - | - | + | - | - | - |
| Quercus dentata Thunb. | 7 | - | + | - | - | - | - |
| Rhamnus ussuriensis Ja. Vassil. | 23 | - | - | - | - | + | - |
| Rhododendron micranthum Turcz. | 119 | + | - | - | - | - | - |
| Ribes latifolium Jancz. | 101 | - | - | - | - | + | - |
| Robinia luxurians (Dieck) C. K. Schneid. | 8 | - | + | - | - | - | - |
| Rosa gallica L. ‘Scharlachglut’ | 57 | + | - | - | - | - | - |
| Rosa kamtschatica Vent. | 1 | - | - | - | - | + | - |
| Salix integra Thunb. ‘Pendula’ | 119 | - | - | - | - | + | - |
| Salix matsudana Koidz. ‘Tortuosa’ | 91 | - | + | - | - | - | - |
| Salix purpurea L. ‘Gracilis’ | 5 | - | - | + | - | - | - |
| Sambucus williamsii Hance | 131 | + | - | - | - | - | - |
| Schisandra grandiflora (Wall.) Hook. et Thoms. var. rubriflora (Rehd. et Wils.) C. K. Schneid. | 127 | - | - | + | - | - | - |
| Spiraea mongolica Maxim. | 107 | - | - | - | - | + | - |
| Weigela decora (Nakai) Nakai | 101 | + | - | - | - | - | - |
| Weigela decora (Nakai) Nakai | 91 | + | - | - | - | - | - |
| Всего: 57 | 11 | 12 | 14 | 5 | 11 | 4 |
В поздне-холодные годы 21 века вымерзли полностью и удалены из коллекции 57 экземпляров, от 14 после зимы 2005/06 гг. до 4 - после зимы 2012/13 гг. Теперь посмотрим, вымерзали ли и сколько деревьев и кустарников в ранне-тёплые годы (таблица 7).
Таблица 7. Древесные растения, вымерзшие в Ботаническом саду Петра Великого в ранне-тёплые годы 21 века
| Название растений | Участок | 2007 | 2008 | 2014 | 2015 | 2016 | 2019 | 2020 |
| Acer negundo L. subsp. californicum (Torrey et Gray) Wesmael | 69 | + | - | - | - | - | - | - |
| Acer opalus Mill. | 19 | - | - | + | - | - | - | - |
| Alnus incana (L.) Moench f. pinnatifida Wahlenb. | 90 | - | - | - | + | - | - | - |
| Aucuba albopunctifolia E. T. Wang | 98 | - | - | - | - | - | - | + |
| Aucuba japonica Thunb. ‘Variegata’ | 99 | - | - | - | - | - | - | + |
| Berberis bretschneideri Rehder | 11 | - | - | + | - | - | - | - |
| Berberis buxifolia Lam. | 99 | - | - | - | - | + | - | - |
| Berberis x ottawensis C. K. Schneid. ‘Purpurea’ | 77 | + | - | - | - | - | - | - |
| Cotoneaster divaricatus Rehder et Wils. | 49 | + | - | - | - | - | - | - |
| Cotoneaster foveolatus Rehder et Wils. | 49 | + | - | - | - | - | - | - |
| Crataegus maximowiczii C. K. Schneid. | 85 | + | - | - | - | - | - | - |
| Dipteronia sinensis Oliv. | 101 | - | - | - | - | + | - | - |
| Erica carnea L. | 130 | - | - | - | - | + | - | - |
| Genista suanica Schischk. | 99 | - | - | - | - | + | - | - |
| Genista tinctoria L. ‘Royal Gold’ | 91 | - | - | - | + | - | - | - |
| Hedera pastuchovii Woronow | 126 | - | - | - | - | + | - | - |
| Hydrangea petiolaris Siebold et Zucc. | 71 | + | - | - | - | - | - | - |
| Idesia polycarpa Maxim. | 99 | - | - | - | - | + | - | - |
| Ilex rugosa Fr. Schmidt | 98 | - | - | - | - | - | - | + |
| Kolkwitzia amabilis Graebn. | 108 | - | + | - | - | - | - | - |
| Laburnum anagyroides Medik. ‘Quercifolium’ | 57 | + | - | - | - | - | - | - |
| Leycesteria formosa Wall. | 99 | - | - | - | + | - | - | - |
| Lonicera ruprechtiana Regel | 118 | - | - | - | - | + | - | - |
| Louiseania ulmifolia (Franch.) Pachom. | 140 | - | - | - | - | - | - | + |
| Padus maackii (Rupr.) Kom. | 36 | + | - | - | - | - | - | - |
| Phlomis fruticosa L. | 99 | - | - | - | - | - | + | - |
| Picea orientalis (L.) Peterm. | 77 | + | - | - | - | - | - | - |
| Platycladus orientalis (L.) Franco | 77 | - | + | - | - | - | - | - |
| Platycrater arguta Siebold et Zucc. | 99 | - | - | - | + | - | - | - |
| Platycrater arguta Siebold et Zucc. | 98 | - | - | - | - | - | - | + |
| Prinsepia sinensis (Oliv.) Bean | 130 | - | - | - | - | + | - | - |
| Rhamnus alpina L. | 48 | + | - | - | - | - | - | - |
| Rhamnus alpina L. | 8 | - | - | + | - | - | - | - |
| Rhodotypos scandens (Thunb.) Makino | 91 | - | - | - | + | - | - | - |
| Ribes aureum Pursh | 18 | - | - | + | - | - | - | - |
| Rosa multiflora Thunb. | 140 | + | - | - | - | - | - | - |
| Ruta graeolens L. | 98 | - | - | - | - | - | + | - |
| Salix schwerinii E. Wolf | 34 | - | + | - | - | - | - | - |
| Sambucus canadensis L. ‘Acutiloba’ | 132 | + | - | - | - | - | - | - |
| Staphylea colchica Stev. | 107 | - | - | - | - | - | + | - |
| Vitis labrusca L. | 91 | - | - | - | - | + | - | - |
| Vitis riparia Michx. ‘Severnij Belij’ | 113 | - | - | - | - | - | + | - |
| Всего: 42 | - | 12 | 3 | 4 | 5 | 9 | 4 | 5 |
Во втором случае число погибших растений в ранне-тёплые годы меньше - 42 шт., от 12 шт. после зимы 2006/07 г. до 3 шт. в следующем 2008 г. Тенденция к уменьшению числа вымерзших растений в ранне-тёплые годы по сравнению с поздне-холодными наблюдается. Хотя деревья и кустарники могут вымерзать при любой метеорологической ситуации, и в нормальные, и даже в мягкие зимы.
Следует иметь в виду, что в последние годы появляется всё более случаев гибели растений не от вымерзания, а от болезней - корневых гнилей и гнилей ствола, связанных с фитофторой и грибами-базидиомицетами, и другими специфическими возбудителями.
Биоклиматическая цикличность имеет место на фоне изменений климата. Но условия существования растений в ранне-тёплые и поздне-холодные годы меняются. Если посмотреть этот 42-летний ряд, он заметно неоднороден. Если в начале 1980-х гг. преобладали поздне-холодные годы с самыми жёсткими условиями для перезимовки растений, то во втором-третьем десятилетиях 21 в. (после 2007 г.) преобладают ранне-тёплые годы. А у поздне-холодных (2011, 2012, 2013 гг.) термическая характеристика совсем другая, чем в начале этого ряда.
С потеплением климата и повышением теплообеспеченности улучшаются условия перезимовки, уменьшается повреждаемость древесных растений морозами. Но одновременно это может способствовать и провоцировать распространение болезней и вредителей, создавать более лучшие условия для смещения границ многих возбудителей и переносчиков болезней и продвижения их на север. Так, например, в случае гибели деревьев она часто объясняется абиотическими факторами или физиологическими причинами — вымоканием, засухой, обморожением, воздействием высоких и слишком низких температур, загрязнением воздуха, кислотными дождями, нехваткой микро- и макроэлементов, и т.п. А на самом деле, зачастую, это действие почвообитающих фитофтор, агентов невидимой смерти. Подсчитано, что в 66 % всех болезней тонких корней и более 90 % гнилей корневой шейки виноваты фитофторы. Дело усугубляется тем, что эти фитопатогены обычно не обнаруживаются, что приводит к неверным диагнозам и, следовательно, к накоплению и распространению инфекции. Чаще всего истинные причины, то есть фитофторы, маскируются вторичными паразитами или даже сапротрофами. Фитофторы в виде мельчайших покоящихся структур хламидоспор и ооспор (20–50 мкм), попадают с растительными остатками в почву, где могут сохраняться от 3 до 20 лет и более. При благоприятных условиях—подходящей влажности, почвенной температуре выше +10° и наличии чувствительного растения, покоящиеся структуры прорастают, образуя либо сразу зооспорангии, либо мицелий—вегетативное тело, на котором уже формируется множество органов бесполого размножения — зооспорангии. Происходящие изменения климата с повышением температуры воздуха и почвы, увеличением осадков и повышением влажности, резким увеличением флуктуаций климатических факторов неизбежно воздействуют и на активность, жизненный цикл и сохранение патогенных организмов (Веденяпина, Фирсов, 2014; Веденяпина и др., 2014 а, б).
Зима 1986/87 г. была последней в 20 веке, которая вызвала столь массовые обмерзания древесных растений в Санкт-Петербурге. Летом по шкале П. И. Лапина нами были зафиксированы результаты обмерзания более 1000 видов и форм древесных растений (Фирсов, Фадеева, 2009). Основная особенность той зимы - рекордно низкие температуры января. Температура воздуха стала неуклонно падать, в среднем на 2,1о в сутки, понизившись за 11 сут. более чем на 23о. Десятого января 1987 г. среднесуточная температура достигла рекордной отметки -33,6о. Критические морозы, когда среднесуточная температура воздуха превышала -30о, держались в течение четырёх суток. Минимальная температура 10 января (-34,7о) - самая холодная во второй половине 20 века. 19 видов погибли в течение этой зимы (Abies nordmanniana и др.). Ещё у 21 вида отмечалась гибель части экземпляров (Morus alba, Juglans regia и др.). Кроме погибших, более чем у половины таксонов парка имело место в той или иной степени обмерзание побегов старше одного года. У многих из них обмёрзли скелетные ветви, надземная часть до уровня снега или корневой шейки, 40 из них погибли на следующий год, выпад продолжался и в последующие годы. Так, Vitis candicans был получен черенками в 1956 г., в 1987 г. он обмёрз до корневой шейки и окончательно погиб на следующий год в возрасте 32 лет (таблица 8). В таблице 8 приводятся результаты обмерзания деревьев и кустарников в парке БИН в поздне-холодные (1987 и 2013 г.) и ранне-тёплые (2019 и 2020) годы в баллах по шкале П. И. Лапина. Индекс «с» означает, что обмёрзли скелетные ветви (то есть значительная часть кроны).
Таблица 8. Обмерзание древесных растений в Ботаническом саду Петра Великого в поздне-холодные и ранне-тёплые годы
| Название растений | Годы ПХ | Годы РТ | ||
| 1987 | 2013 | 2019 | 2020 | |
| Abies firma Siebold et Zucc. | 4c | 1 | 1 | 1 |
| Abies nordmanniana (Stev.) Spach | 7 | 1 | 1 | 1 |
| Acer carpinifolium Siebold et Zucc. | 6 | 2 | 1 | 1 |
| Acer cissifolium Siebold et Zucc. | 7 | 1 | 1 | 1 |
| Acer hyrcanum Fisch. et C. A. Mey. | 4-7 | 1 | 1 | 1 |
| Acer opalus Mill. | 4c-6 | 1 | 1 | 1 |
| Actinidia arguta (Siebold et Zucc.) Planch. ex Miq. | 4c-7 | 2 | 2 | 2 |
| Amorpha fruticosa L. | 6 | 4с | 2 | 2 |
| Aralia elata (Miq.) Seem. | 4-7 | 1 | 1 | 1 |
| Armeniaca vulgaris Lam. | 7 | 2 | 2 | 1 |
| Carpinus caroliniana Walt. | 4c-5 | 4с | 2 | 1 |
| Carpinus cordata Blume | 4c | 1 | 1 | 1 |
| Catalpa speciosa (Warder ex Barney) Engelm. | 4-5 | 2 | 2 | 2 |
| Cerasus sachalinensis (Fr. Schmidt) Kom. | 4c | 1 | 1 | 1 |
| Chionanthus virginicus L. | 4c | 1 | 1 | 1 |
| Cydonia oblonga Mill. | 4с | 2 | 1 | 1 |
| Deutzia amurensis (Regel) Airy-Shaw | 4-7 | 1 | 1 | 1 |
| Eleutherococcus henryi Oliv. | 5-7 | 2 | 2 | 1 |
| Fagus orientalis Lipsky | 4c-5 | 1 | 1 | 1 |
| Fagus sylvatica L. | 4-4c | 1 | 1 | 1 |
| Forsythia x intermedia Zabel | 5 | 4 | 1 | 1 |
| Fraxinus rhynchophylla Hance | 4c-6 | 1 | 1 | 1 |
| Gymnocladus dioicus (L.) C. Koch | 6 | 2 | 2 | 2 |
| Juglans regia L. | 4c-7 | 1 | 1 | 1 |
| Laburnum alpinum (Mill.) Berchtold et Presl | 4c | 1 | 1 | 1 |
| Ligustrum ovalifolium Hassk. | 5 | 2 | 2 | 2 |
| Malus sieboldii (Regel) Rehder | 6 | 2 | 1 | 1 |
| Metasequoia glyptostroboides Hu et W. C. Cheng | 4-7 | 2-3 | 2 | 2 |
| Morus alba L. | 4-7 | 2-4 | 1-2 | 1 |
| Oplopanax elatus (Nakai) Nakai | 7 | 1 | 1 | 1 |
| Padellus mahaleb (L.) Vass. | 4c-5 | 2 | 1 | 1 |
| Pinus funebris Kom. | 4 | 1 | 1 | 1 |
| Prinsepia sinensis (Oliv.) Bean | 4c-6 | 2 | 2 | 2 |
| Robinia pseudoacacia L. | 4c-7 | 2 | 2 | 2 |
| Sambucus nigra L. | 4c-6 | 2 | 2 | 2 |
| Weigela hortensis (Siebold et Zucc.) C. Koch | 6 | 4c | 2 | 2 |
Год 2013, так же как и 1987, относится к поздне-холодным. Но термические условия его уже другие. Абсолютный минимум температуры воздуха зимой 2012/13 г. достигал только -22,3о. А температура января 2013 г. составила всего -6,1о, в отличие от января 1987 г., когда онаопускалась до -17,9о. И повреждения зимними морозами были заметно меньше чем в 1987 г. Тем не менее, в целом существует и до сих пор сохраняется различие между обмерзанием деревьев и кустарников в поздне- холодные и ранне-тёплые годы. В случае холодных и аномально суровых зим, которые обычно и приходятся на поздне-холодные годы, различие становится очень заметным. В других случаях оно нивелируется, но в целом не исчезает. При этом в ранне-тёплые годы большинство видов вообще перестают обмерзать (балл 1) или у них балл обмерзания не более 2. При этом существуют виды, преимущественно кустарники более южного происхождения, такие как вейгелы, с длительно растущими побегами, которые могут не вызревать до осени и уходят в зиму с растущими, не одревесневшими концами побегов. Такие растения обычно обмерзают ежегодно, невзирая на особенности вегетационного сезона и характеристику зимних условий. В целом в ранне-тёплые зимы подавляющее большинство видов парка имеют балл обмерзания 1-2. И лишь у единичных особей обмерзают побеги старше одного года (если это не относится к усыханию от фитофтор или других болезней).
В начале 1970-х гг. фенологи и климатологи обратили внимание на тенденцию к потеплению климата Санкт-Петербурга (Булыгин, Довгулевич, 1974). Заметное потепление началось с 1989 г., который стал самым тёплым (7,6о) в истории на тот период времени. В XXI веке потепление климата усилилось, особенно после 2007 г. Зима 2006/ 07 г. была рекордно короткой и продолжалась лишь 41 день, зато осень длилась почти 5 месяцев (Фирсов и др., 2008). Очень тёплым было лето 2010 г., при рекордно высоких температурах июля (24,4о). Во втором десятилетии XXI века до конца календарного года зима в отдельные года так и не наступает – отсутствует снежный покров, а некоторые растения продолжают вегетацию. Очевидна тенденция к повышению теплообеспеченности и при сравнении среднегодовой температуры воздуха. Она повысилась на 2,5о по отношению к норме климата 20 столетия, что можно считать очень значительным потеплением. Год 2015 стал самым тёплым за период наблюдений с середины 18 века: 7,7о (его позже превзошёл 2020 г.: 8,3о). Заметно потеплела и минимальная температура воздуха (Фирсов, Фадеева, 2020). С потеплением климата в начале XXI века в Санкт-Петербурге заметно увеличилось число видов деревьев и кустарников, достигших репродуктивного состояния. У целого ряда видов впервые за длительный период интродукции было получено семенное потомство: Acer japonicum Thunb. (Волчанская и др., 2010); Picea glehnii (F. Schmidt) Mast. (Фирсов и др., 2015); Fraxinus oxycarpa M. Bieb. ex Willd.; Fraxinus pojarkoviana V. Vassil.; Сydonia oblonga Mill. (); Carpinus orientalis Mill. (Фирсов, Ткаченко, 2018); Acer capillipes Maxim. (Фирсов и др., 2018); Pyrus zangezuraFirsov G.A. et al., 2017 Maleev (Ткаченко и др., 2019); Aristolochia macrophylla Lam. (Ткаченко и др., 2020). У некоторых видов отмечено первое цветение, как, например, у Decaisnea fargesii Franch. (Фирсов, 2019).
В таблице 9 приводится список видов, достигших фазы плодоношения в начале третьего тысячелетия, в первые два десятилетия 21 века. При этом многие из них десятки лет до этого находились в вегетативном состоянии. В графе 2 приводится год появления всходов. В графе 3 - год первого плодоношения, в графе 4 обозначен тип БКЦ (ранне-тёплый или поздне-холодный), когда впервые наблюдалось плодоношение (у некоторых видов первое цветение).
В таблицу 9 включены 37 видов. И только в 5 случаях начало плодоношения пришлось на поздне-холодный год. В остальных 86,5 % случаев достижения репродуктивного состояния имели место в ранне-тёплые годы. Это имеет явно положительный эффект. По мнению Н. И. Булыгина, оценивать устойчивость тех или иных видов деревьев и кустарников и перспективность для разведения следует в поздне-холодные годы. А работы, связанные с размножением растений, проводить в ранне-тёплые биоклиматические циклы. Из данных таблицы 9 видно, что ранне-тёплые годы полезны для репродуктивной сферы растений. Однако, как изменяются качества семян и другие характеристики (качество пыльцы, обилие плодоношения и т.д.) в ранне-тёплые и поздне-холодные годы (есть ли различие и каково оно?) - вопрос пока что не изученный, особенно на фоне изменений климата. Известно, что на будущий урожай и качество семян влияет не только уровень температуры, но и особенности погоды во время цветения. Прежде всего осадки, их количество и продолжительность. А для ветроопыляемых растений может иметь значение и ветровой режим. Поэтому следует проводить дальнейший непрерывный мониторинг. И в своей работе учитывать такое явление, как биоклиматическую цикличность.
Таблица 9. Деревья и кустарники Ботанического сада Петра Великого, достигшие репродуктивного состояния в 2007-2020 г.
| Название растений | Год посева семян | Год первого плодоношения (или цветения (Цв)) | Поздне-холодный или ранне-теплый год (ПХ или РТ) |
| 1 | 2 | 3 | 4 |
| Abies alba Mill. | 1971 | 2013 | ПХ |
| Abies arizonica Merr. | 1973 | 2015 | РТ |
| Abies gracilis Kom. | 1986 | 2015 | РТ |
| Acer capillipes Maxim. | ~1995 | 2014 | РТ |
| Acer cissifolium (Siebold et Zucc.) C. Koch | 1999 | 2019 | РТ |
| Acer griseum Pax | 1999 | 2019 | РТ |
| Acer grosseri Pax | 2006 | 2019 | РТ |
| Acer henryi Pax | 2001 | 2013 | ПХ |
| Acer japonicum Thunb. ex Marray | 1999 | 2015 | РТ |
| Acer mayrii Schwer. | 2004 | 2019 | РТ |
| Acer opalus Mill. | 1989 | 2019 | РТ |
| Acer palmatum Thunb. ex Murray | 1981 | 2007 | РТ |
| Acer pensylvanicum L. | ~2001 | 2015 (Цв.) | РТ |
| Acer saccharum Marshall subsp. nigrum (Michx. f.) Desmarais | 1977 | 2011 | ПХ |
| Acer velutinum Boiss. | ~2008 | 2015 | РТ |
| Betula medvedievii Regel | 1986 | 2015 | РТ |
| Callicarpa japonica Thunb. | 2009 | 2015 (Цв.) | РТ |
| Catalpa bungei C. A. Mey. | 1997 | 2007 | РТ |
| Chosenia arbutifolia (Pall.) A. Skvorts. | ~1998 | 2019 | РТ |
| Cladrastis kentukea (Dum.-Cours.) Rudd | 1994 | 2013 | ПХ |
| Cornus walteri Wanger. | 2019 | РТ | |
| Corylopsis sinensis Hemsl. | 2002 | 2015 (Цв.) | РТ |
| Corylus colurna L. | ~1980 | 2015 | РТ |
| Crataegus pinnatifida Bunge | ~1985 | 2015 | РТ |
| Cydonia oblonga Mill. | 1949 | 2014 | РТ |
| Euonymus latifolius (L.) MIll. | 2011 | 2020 | РТ |
| Fraxinus oxycarpa Willd. | ~1965 | 2014 | РТ |
| Fraxinus pojarkovianae V. Vasssil. | 1980 | 2015 | РТ |
| Hydrangea robusta Hook. et Thoms. | 1996 | 2014 | РТ |
| Itea virginica L. | ~2007 | 2015 (Цв.) | РТ |
| Liriodendron tulipifera L. | 1956 | 2019 | РТ |
| Ostrya carpinifolia Scop. | 1999 | 2019 | РТ |
| Pinus pallasiana D. Don | 1960 | 2007 | РТ |
| Pterocarya stenoptera DC. | 1988 | 2011 | ПХ |
| Pyrus zangezura Maleev | 2003 | 2016 | РТ |
| Rhododendron brachycarpum D. Don subsp. tigerstedtii Nitzelius | 2003 | 2019 | РТ |
| Rhus typhina L. | ~2013 | 2020 | РТ |
Изменение климата в сторону потепления дает возможность выращивать большее число видов из семян местной репродукции и, в конечном счете, способствовать их акклиматизации и введению в практику городского озеленения. Почему же происходит такой «всплеск» репродуктивных способностей растений, как это видно из данных таблицы 9?
– Этому способствует смягчение зимних температур. Повышение минимальной температуры воздуха уже привело к смещению границ зимней устойчивости древесных растений, и границы зон всё сильнее смещаются к северу и востоку (Фирсов, Хмарик, 2016). Особенно важно повышение зимних температур для тех видов деревьев и кустарников, у которых цветочные почки закладывают летом года, предшествующего цветению. В прошлом у таких видов в холодные зимы они просто вымерзали.
– Значительное удлинение вегетационного сезона (Фадеева, Фирсов, 2010) способствует вызреванию вегетативных побегов, а также плодов и семян. В прошлом в неблагоприятные годы не вызревали семена даже у видов местной флоры (как у Acer platanoides L. в 1976 г.).
– Очень важное увеличение безморозного периода, особенно для раннецветущих видов, таких, как Acer saccharinum L. (Булыгин, Фирсов, 1985).
Для ряда видов для их развития репродуктивной сферы (например, с востока Северной Америки, как Carya ovata (Mill.) K. Koch) важное значение имеют высокие летние температуры. После аномально жаркого лета 2010 г., с так называемым блокирующим антициклоном, на научно-опытной станции БИН РАН «Отрадное» отмечен самосев Cercidiphyllum japonicum Siebold et Zucc. (Фирсов, Хмарик, Трофимук, 2020).
– Возможно, что для насекомоопыляемых видов более высокие температуры воздуха в период цветения оказывают благоприятное влияние на насекомых-опылителей и на возможность опыления.
– Такому «всплеску» способствует и общее увеличение видов культивируемой дендрофлоры. Если после Великой Отечественной войны и в 1980-е гг. коллекция парка-дендрария Ботанического сада Петра Великого Ботанического института имени В. Л. Комарова насчитывала 600-800 видов и форм, то сейчас она превышает 1200 таксонов. Потепление климата значительно расширяет возможности привлечения в культуру многих теплолюбивых видов более южных широт (Ткаченко и др., 2020).
На фоне продолжающегося потепления климата обнаружен самосев целого ряда видов, у которых он ранее не наблюдался (Carpinus betulus L., Chamaecyparis pisifera (Siebold & Zucc.) Endl., Cerasus maximowiczii (Rupr.) Kom. & Aliss., Spiraea betulifolia Pall. и многие другие), что является важным показателем адаптации и признаком возможной будущей натурализации вида на этой территории. При этом многие виды могут стать и становятся потенциально инвазионными (Фирсов и др., 2017), что ставит вопросы об их тщательном мониторинге и культуре в контролируемых условиях. Особенности появления и развития самосева у разных видов в ранне-тёплые и поздне-холодные годы - вопрос дальнейших углублённых исследований. При этом надо учитывать, что цикличность существует при любом климате. И интродукторы, садоводы и лесоводы должны учитывать её в своей работе.
На фоне выраженного потепления климата имеет место чередование ранне-тёплых и поздне-холодных биоклиматических циклов или отдельных лет. Когда одни и те же растения обнаруживают совсем разные показатели фенологического биоритма, уровни адаптированности и репродуктивной способности.
Заключение
В Ботаническом саду Петра Великого БИН РАН непрерывный фенологический мониторинг по программе Календаря природы ведётся постоянно с 1980 г. Анализ сроков наступления и продолжительности сезонов за 42 года наблюдений (1980-2021 гг.) показал, что на фоне потепления климата биоклиматическая цикличность проявляется в чередовании ранне-тёплых (1989, 1990, 1992, 1995, 2007, 2008, 2014, 2015, 2016, 2019, 2020 гг.) и поздне-холодных (1980, 1982, 1985, 1987, 1996, 1998, 2003, 2004, 2006, 2011, 2012, 2013 гг.) лет. Это отражается на зимостойкости и репродуктивной способности древесных растений. Все наиболее суровые и неблагоприятные зимы пришлись на поздне-холодные годы. В ранне-тёплые годы по сравнению с поздне-холодными заметно теплее в самые холодные месяцы года (в феврале на 7,2о, январе на 4,4о). Достаточно заметное различие проявляется и весной, по июнь включительно. Этим создаются лучшие условия для перезимовки растений. В ранне-тёплые годы продолжительность безморозного периода увеличивается на 9 сут. Сокращается продолжительность зимы на 52 сут., за счёт этого удлиняются весенний и летний сезоны.
Весь опыт разведения древесных растений в Санкт-Петербурге на протяжении трёх веков показывает, что здесь основным фактором их выживания являются аномально суровые, или критические зимы. После нормальных и теплых зим у большинства видов деревьев и кустарников, культивируемых в ботанических садах Санкт-Петербурга, обмерзания отсутствуют или не превышают концов годичных побегов. Если последствия теплых зим до последнего времени сказывались в основном на цветении и плодоношении, то после аномально суровых зим, как правило, наблюдается массовая гибель многих видов или сильные обмерзания скелетных ветвей. Все такие зимы за период 1955-2009 гг. приурочены к поздне-холодным биоклиматическим циклам. Небольшой возраст многих деревьев и кустарников, выращиваемых в ботанических садах Санкт-Петербурга, есть результат того, что эти виды неоднократно вводились в культуру в разные периоды времени, и даже могли цвести и плодоносить, но через некоторое время погибали после суровых зим. Многие теплолюбивые экзоты могут существовать здесь только в промежутке между аномально суровыми зимами. В некоторых случаях культура таких растений оправдана. Но для уличного озеленения и массового разведения пригодны лишь наиболее зимостойкие деревья и кустарники. Вот почему особую научную ценность имеет многолетний мониторинг древесных растений в ботанических садах в годы с разной метеорологической ситуацией и в разные биоклиматические циклы. С потеплением климата за последние десятилетия повторяемость критических зим уменьшается, и после аномально-холодной зимы 1986-87 г. подобных зим в Санкт-Петербурге не повторялось, хотя это не исключает вероятности их наступления в будущем.
До сих пор ранне-тёплые годы рассматривались в качестве благоприятной биоклиматической ситуации. Однако, если потепление климата будет продолжаться, то ситуация может измениться и оптимальными для роста и развития древесных растений станут, наоборот, поздне-холодные годы. Поэтому сейчас резко возрастает роль фенологических наблюдений и значение непрерывных рядов дендрофеноиндикаторов.
В деятельности ботанических садов и других интродукционных центров нельзя обойтись без фенологических наблюдений, что позволит выделить феноритмотипы и обрабатывать накопленные ряды. Такие методы исследований, хотя они являются визуальными и относительно простыми, позволяют охватить наблюдениями и сделать оценку для всех или большей части коллекционных фондов. Особую ценность имеют непрерывные ряды наблюдений длиннее 10-15 лет.
Благодарности
Работа выполнена в рамках государственного задания по плановой теме «Коллекции живых растений Ботанического института им. В. Л. Комарова (история, современное состояние, перспективы использования)», номер АААА-А18-118032890141–4.
Литература
Афанасьев С. Л. Возможные причины пульсации и цикличности геологических процессов // Ритмика природных явлений: Тезисы докладов к III совещанию, Ленинград, 30 ноября 1976 г. Л.: Географическое общество СССР, 1976. С. 27—30.
Булыгин Н. Е., Довгулевич З. Н. Некоторые результаты математического анализа вековых фенологических рядов // Межвуз. сб. законч. науч. исслед. работ. Вып. 2. Л.: ЛТА, 1974 а. С. 36—40.
Булыгин Н. Е., Довгулевич З. Н. О фенологической тенденции и цикличности в «вековых» фенологических рядах на Северо-Западе России // Лесоводство, лесные культуры и почвоведение. Вып. 3: Межвуз. сб. науч. тр. Л.: ЛТА, 1974 б. С. 25—33.
Булыгин Н. Е. Исследование тенденции и цикличности в фенологических рядах в связи с фитофенологическим прогнозированием на Северо-Западе России // Физико-географич. районир. и прогнозир. Л.: Географич. общ-во СССР, 1975 а. С. 32—36.
Булыгин Н. Е. Тенденция и цикличность в вековых фенологических рядах // Тезисы 12 Международ. ботан. конгресса. Л.: Наука, 1975 б. С. 179.
Булыгин Н. Е., Топпер С. М. Сезонная сопряженность фитофеноиндикаторов на фоне циклических колебаний климата // Ритмика природных явлений: Тезисы докладов к III совещанию, Ленинград, 30 ноября 1976 г. Л.: Географическое общество СССР, 1976. С. 144—147.
Булыгин Н. Е., Бердникова С. В., Михалкина Л. Г. Цикличность в фитоценологических рядах и ее индикационное значение // Лесное хоз-во, лесная, деревообр. и целлюлоз.-бум. пром. Вып. 4.: Межвуз. сборник статей о законч. науч.-исслед. работах. Л.: ЛТА, 1976. С. 8—10.
Булыгин Н. Е., Топпер С. М. Сравнительная эффективность разных методов фенопрогнозов и их использование в проверке массовой фенологической информации // Докл. IV межведомств. совещ. по фенологич. прогнозир. Л.: Географ. общ-во СССР, 1977. С. 13—17.
Булыгин Н. Е. Фенологические наблюдения над древесными растениями. Л.: ЛТА, 1979. 97 с.
Булыгин Н. Е. Сезонно-ритмическая структура годичного цикла развития ландшафта, принципы ее индикации и прогностическое значение // Моделирование и прогнозирование в индикационной дендрофенологии. Л.: ЛТА, 1980. C. 2—44. Деп. в ВИНИТИ, № 1033-81 Деп.
Булыгин Н. Е., Курочкина В. А., Михалкина Л. Г. Исследование корреляционных связей между дендрофеноиндикаторами и последующей теплообеспеченностью вегетационного сезона // Моделирование и прогнозирование в индикационной дендрофенологии. Л.: ЛТА, 1980. С. 62—68. Деп. в ВИНИТИ, № 1033-81 Деп.
Булыгин Н. Е. Биоклиматическая цикличность и зимостойкость древесных интродуцентов // Тез. докл. VIII Дендрол. Конгресса соц. стран. Тбилиси, 1982а. С. 219.
Булыгин Н. Е. Биологические основы дендрофенологии. Л.: ЛТА, 1982б. 80 с.
Булыгин Н. Е., Фирсов Г. А. Клен серебристый в Ленинграде и перспективы его использования в озеленении на Северо-Западе РСФСР. Л.: ЛТА, 1985. 31 с. Деп. в ВИНИТИ, № 6296-85 Деп.
Булыгин Н. Е. Полтора века фенологического мониторинга в Лесном // Известия Санкт-Петербургской Лесотехнической академии. Вып. 4 (162). СПб., 1996а. С. 166—175.
Булыгин Н. Е. Короткопериодные колебания климата и интродукция растений // Труды первой Всероссийской конференции по ботаническому ресурсоведению, 25-30 ноября 1996 года. СПб., 1996б. С. 113.
Булыгин Н. Е., Фирсов Г. А. Биоклиматическая цикличность и адаптация древесных растений муссонного климата при интродукции их на Северо-Запад России // Материалы Международ. конф., посвящ. 50-летию Бот. сада – ин-та ДВО РАН. Владивосток: Дальнаука, 1998. С. 157—160.
Веденяпина Е. Г., Фирсов Г. А. Невидимые фитофторы // Питомник и частный сад. 2014. № 2. С. 40—45.
Веденяпина Е. Г., Волчанская А. В., Малышева В. Ф., Малышева Е. Ф., Фирсов Г. А. Почвообитающие виды рода Phytophtora в Ботаническом саду БИН РАН. I. Первые находки Ph. citricola, Ph. plurivora и Ph. quercina в России // Микология и фитопатология. 2014а. Т. 48. Вып. 4. С. 263—273.
Веденяпина Е. Г., Фирсов Г. А., Волчанская А. В., Воробьев Н. И. Почвообитающие виды рода Phytophthora в Ботаническом саду БИН РАН. II. Результаты двухлетнего мониторинга // Микология и фитопатология. 2014б. Т. 48. Вып. 5. С. 322—332.
Волчанская А. В., Фирсов Г. А., Лаврентьев Н. В. Клён японский (Acer japonicum Thunb.) в Санкт-Петербурге // Вестник ОрелГАУ. 2010. № 2 (23). С. 66—72.
Давитая Ф. Ф. Прогноз обеспеченности теплом и некоторые проблемы сезонного развития природы. Л.: Гидрометеоиздат, 1964. 132 с.
Лапин П. И. Сезонный ритм развития древесных растений и его значение для интродукции // Бюлл. ГБС. 1967. Вып. 65. С. 13—18.
Максимов Е. В. Ритмичность природных явлений // Ритмика природных явлений: Тезисы докладов к III совещанию, Ленинград, 30 ноября 1976 г. Л.: Географическое общество СССР, 1976. С. 5—8.
Ткаченко К. Г., Фирсов Г. А., Яндовка Л. Ф., Волчанская А. В., Староверов Н. Е., Грязнов А. Ю. Груша зангезурская (Pyrus zangezura, Rosaceae) в Санкт-Петербурге // Труды по прикладной ботанике, генетике и селекции. 2019. Т. 180. Вып. 3. C. 12—18.
Ткаченко К. Г., Фирсов Г. А., Волчанская А. В. Качество семян Aristolochia macrophylla Lam. и A. manshuriensis Kom. в Санкт-Петербурге // Труды по прикладной ботанике, генетике и селекции. 2020. Т. 181. № 2. С. 14—22.
Оль А. И. Ритмичность воздействия солнечной активности на ландшафтную оболочку Земли // Ритмика природных явлений: Тезисы докладов к III совещанию, Ленинград, 30 ноября 1976 г. Л.: Географическое общество СССР, 1976. С. 15—17.
Ржонскицкий В. Б. Приливы на Солнце и их влияние на геофизические процессы // Ритмика природных явлений: Тезисы докладов к III совещанию, Ленинград, 30 ноября 1976 г. Л.: Географическое общество СССР, 1976. С. 23—27.
Фадеева И. В., Фирсов Г. А. Индикационное значение дендрофонологического ряда зацветания Alnus incana в феностационаре Санкт-Петербургской лесотехнической академии // Дендрология в начале XXI века: Сборник материалов Международных научных чтений памяти Э. Л. Вольфа, 6-7 октября 2010 года, Санкт-Петербургская государственная лесотехническая академия им. С. М. Кирова. СПб.: Изд-во Политехн. ун-та. 2010. C. 210—214.
Фирсов Г. А. К проблеме дендрологического районирования территории Северо-Запада России // Бюлл. ГБС. Вып. 185. 2003. С. 3-8.
Фирсов Г. А. Памяти Николая Евгеньевича Булыгина (12 VIII 1924 – 22 V 2002) // Бот. журн. 2004. Т. 89. № 3. С. 186—190.
Фирсов Г. А., Бялт В. В. Обзор древесных экзотов, дающих самосев в г. Санкт-Петербурге (Россия) // Российский журнал биологических инвазий. 2015. № 4. С. 129—152.
Фирсов Г. А., Ткаченко К. Г., Лаврентьев Н. В. Инвазионный потенциал Quercus rubra L. в Санкт-Петербурге // Вестник Удмуртского гос. университета. Сер. Биология. Науки о Земле. 2017. Т. 27. Вып. 3. С. 297—305.
Фирсов Г. А., Ярмишко В. Т. Николай Евгеньевич Булыгин как дендролог и фенолог // Бот. журн. 2005. Т. 90. № 4. С. 604—621.
Фирсов Г. А., Фадеева И. В., Волчанская А. В. Влияние метео-фенологической аномалии зимы 2006/07 года на древесные растения в Санкт-Петербурге // Вестник Московского гос. университета леса (Лесной вестник). 2008. № 6 (63). С. 22—28.
Фирсов Г. А., Фадеева И. В. Аномально-суровая зима 1986-87 гг. и зимостойкость древесных растений в Санкт-Петербурге // Научное обозрение. 2009. № 3. С. 8—19.
Фирсов Г. А., Фадеева И. В. Календарь природы Ботанического сада Ботанического института им. В. Л. Комарова РАН // Древесные растения: фундаментальные и прикладные исследования. Вып. 2. М.: ФГБУН ГБС РАН, 2013. C. 111—125.
Фирсов Г. А. Древесные растения ботанического сада Петра Великого (XVIII-XXI вв.) и климат Санкт-Петербурга // Ботаника: история, теория, практика (к 300-летию основания Ботанического института им. В. Л. Комарова Российской академии наук): Труды международной научной конференции. СПб.: Изд-во СПбГЭТУ «ЛЭТИ», 2014. C. 208—215.
Фирсов Г. А., Волчанская А. В., Ткаченко К. Г. Ель Глена (Picea glehnii (F. Schmidt) Mast., Pinaceae) в Санкт-Петербурге // Вестник Удмуртского ун-та. Серия Биология. Науки о Земле. 2016. Т. 26. Вып. 3. С. 58—65.
Фирсов Г. А., Волчанская А. В., Ткаченко К. Г. Клён волосовидный (Acer capillipes Maxim. ex Miq., Sapindaceae) в Санкт-Петербурге // Вестник Воронежского гос. университета. Серия: Химия. Биология. Фармация. 2018. № 1. С. 152—158.
Фирсов Г. А., Ткаченко К. Г. Граб восточный (Carpinus orientalis Mill., Betulaceae) в Санкт-Петербурге // Бюлл. ГБС. 2018. Вып. 204. № 2. С. 9—15.
Фирсов Г. А. Первое цветение Decaisnea fargesii Franch. (Lardizabalaceae) в Ботаническом саду Петра Великого БИН РАН // Цветоводство: история, теория, практика: Сборник статей IX Международной научной конференции (7-13 сентября 2019 г., г. Санкт-Петербург). СПб., 2019. С. 226—228.
Фирсов Г. А., Фадеева И. В. Изменение климата и возможные изменения ассортимента древесных растений Санкт-Петербурга // Бюлл. ГБС. 2020. Вып. 206. № 1. С. 57—63.
Фирсов Г. А., Хмарик А. Г., Трофимук Л. П. Багрянник японский (Cercidiphyllum japonicum Siebold et Zucc.) на северо-востоке Карельского перешейка (Ленинградская область) // Бюлл. ГБС. 2020. № 2. Вып. 206. С. 25—30.
Фирсов Г. А., Волчанская А. В. Древесные растения в условиях климатических изменений в Санкт-Петербурге. Москва: "МАСКА", 2021. 128 с.
Шнитников А. В. Предисловие // Ритмика природных явлений: Тез. докл. к III совещанию, Ленинград, 30 ноября 1976 г. Л., 1976. С. 3—4.
Шульц Г. Э. Фенологические наблюдения - индикатор циклических колебаний климата // Изв. Всесоюзн. географ. общ-ва. 1978. № 6. С. 498—504.
Ярмишко В. Т. Николай Евгеньевич Булыгин. К 75-летию со дня рождения // Бот. журн. 1999. Т. 84. № 12. С. 134—140.
Beckett K. Hardiness Zones and the British Isles // Krussmann G. Manual of Cultivated Conifers. Portland, Oregon: Timber Press, 1995. P. 7—8.
Daly Ch., Widrlechner M. P., Halbleib M. D., Smith J. I., Gibson W. P. Development of a New USDA Plant Hardiness Zone Map for the United States // Journal of applied meteorology and climatology. 2012. Vol. 51. P. 242—264.
Firsov G. A., Volchanskaya A. V., Tkachenko K. G., Staroverov N. E., Gryaznov A. Yu. Cydonia oblonga Mill. (Rosaceae) at the Peter the Great Botanical Garden // Proceedings on applied botany, genetics and breeding. 2016. 177 (4). P. 28—36. https://doi.org/10.30901/2227-8834-2016-4-28-36 .
Magarey R. D., Borchert D. M., Schlegel J. W. Global plant hardiness zones for phytosanitary risk analysis // Scientia Agricola. 2008. Vol. 65. P. 54—59.
Prokhorov A. A., Eglacheva A. V., Andrjusenko V. V., Dementiev P. P., Zarodov A. Yu. A comparative and statistical analysis of Pinopsida in the collections of Russia's botanic gardens, under varying climatic conditions // EuroGard V: Botanic gardens in the age of climate change – supplementary proceedings. Ulmus, 2012. № 14. P. 80—90.
Rehder A. A Manual of Cultivated Trees and Shrubs Hardy in North America. N. Y.: The Macmillan Company, 1949. 996 p.
Warda H. - D. Foreword to the Second Edition // Krussmann G. Manual of Cultivated Conifers. Portland, Oregon: Timber Press, 1995. P. 5.
