

Введение
Интродукция растений мировой флоры позволяет решать одновременно две задачи: сохранение биоразнообразия в природе и расширение ассортимента полезных растений в каждом регионе. Дикорастущие виды по сравнению с культиварами гораздо более устойчивы к неблагоприятным условиям, болезням и вредителям, что обусловливает их перспективность для зеленого строительства. Применяемые при этом агротехнические мероприятия позволяют выращивать ex situ виды, не характерные для природно-климатических условий интродукционного пункта, и за счет этого постоянно увеличивать число новых цветочно-декоративных растений. В этом отношении интересными и перспективными видами являются представители рода Iris L., и одно из заметных мест среди интродуцентов этого родового комплекса занимает I. рseudacorus L. (Ирис ложноаировый, или болотный), результаты комплексных интродукционных исследований которого представлены в данной статье. Работа проводилась в лаборатории цветоводства Донецкого ботанического сада (ДБС) на протяжении 16 лет.
Целью работы является оценка адаптационных возможностей I. рseudacorus в условиях степной зоны на основе комплексных интродукционных исследований. В задачи исследований входят изучение морфологии вегетативных и генеративных органов, феноритмики, способности к репродукции, онтогенеза, а также оценка успешности интродукции с определением путей использования вида в зеленом строительстве региона.
Объекты и методы исследований
В ДБС I. рseudacorus интродуцирован семенами, полученными по делектусному обмену в 2001 г. из Франции. В природе он растет на болотах, по берегам и мелководьям рек и озер Атлантической и Средней Европы, Кавказа, Западной Сибири, Средиземноморья и Малой Азии, при этом корневище обычно погружено в воду, листья и цветоносы находятся над водой (Федченко, 1935; Энциклопедия…, 2017). Тем не менее, при культивировании в засушливых условиях Донбасса этот вид проходит все фазы годичного цикла развития и сохраняет декоративность в течение всего вегетационного периода.
Изучение морфологических особенностей и сезонного ритма развития проводили по общепринятой методике с учетом основных этапов вегетации (Методика…, 1975), феноритмотип определяли по И. В. Борисовой (1972), жизненную форму – в соответствии с классификацией A. Б. Безделева, Т. А. Безделевой (2006) и дополнениями согласно В. В. Тарасову (2012), семенную продуктивность – по И. В. Вайнагий (1974), онтогенез изучали в соответствии с классификацией Т. А. Работнова (1964, 1965), дополненной А. А. Урановым (1975), с использованием методики И. И. Игнатьевой (1983). Определение успешности интродукции проводили согласно шкале В. В. Бакановой (1984).
Результаты и обсуждение
В условиях культуры открытого грунта ДБС I. рseudacorus – многолетний летнезеленый травянистый короткокорневищно-кистекорневой рыхлодерновинный, симподиально нарастающий, вегетативно малоподвижный поликарпик с полурозеточным прямостоячим побегом. Листья светло-зеленые, мечевидные, 2,5–3,2 см шириной, 47–75 см длиной. Цветонос 50–70 см высотой, облиственный, с 2–4 боковыми ветвями, несущими по 2–3 золотисто-желтых цветка около 7 см диаметром, 4 см высотой. Плод – нижняя синкарпная трехгнездная коробочка длиной до 8 см (рис. 1), семена большей частью полукруглые, сплюснутые, кофейно-коричневые (рис. 2).
Корневище плагиотропное, на поперечном срезе красновато-розовое, состоит из сильно утолщенных и укороченных годичных приростов (звеньев) 5–8 см длиной и 2,5–3,5 см в диаметре. Звенья корневища текущего года прикрыты влагалищами листьев и остатками отмерших листьев генерации прошлого года, снизу несут бородку из придаточных корней, ежегодно отмирающих. Плагиотропное кореневище обусловливает рыхлокустовой тип ветвления, но в аридных условиях Донбасса звенья корневища I. рseudacorus сближены, в результате чего растение приближается к плотнокустовому типу. По характеру фенологического развития в годичном цикле I. рseudacorus относится к весенне-летне-осеннезеленым видам поздневесеннего-раннелетнего цветения. Вегетацию начинает после оттаивания почвы, заканчивает в начале ноября. Цветет в мае на протяжении 10 дней (табл. 1).

Рис. 1. Плоды Iris pseudacorus L.
Таблица 1. Основные фенологические фазы Iris pseudacorus L. (2002–2016 гг.).
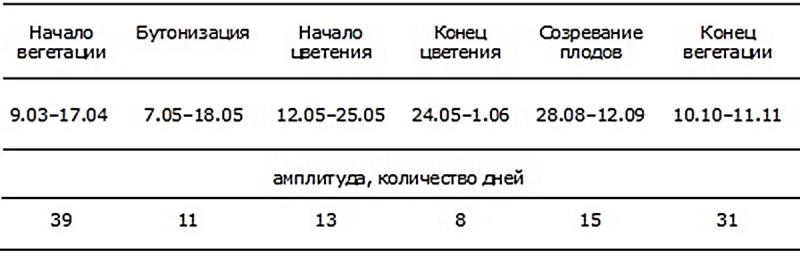
Начало вегетации I. рseudacorus обусловлено термическим фактором в сочетании с фактором временным, а именно показателем среднесуточных температур воздуха в течение определенного периода времени. Таким определяющим показателем мы считаем сумму среднесуточных температур за декаду, предшествующую отмеченной фенодате. Нижним пределом, определяющим начало вегетации I. рseudacorus, является точка устойчивого перехода среднесуточных температур воздуха через отметку +10° С (при этом сумма температур за предыдущую декаду составляет не менее +100° С). Кратковременное повышение температуры воздуха (менее недели), даже значительное, к активизации ростовых процессов не приводит.
Анализ сезонного ритма развития I. рseudacorus показал, что смена фенофаз в их годичном цикле соответствует погодно-климатическому ритму интродукционного пункта: амплитуды основных фенофаз определяются главным образом температурным режимом. Так, сроки прихода весны в разные годы могут колебаться в пределах полутора месяцев – и, соответственно, почти в таких же пределах колеблется дата весеннего отрастания. Бутонизация и цветение происходят в период оптимизации температурных условий – и амплитуды колебания их сроков сокращаются до двух недель. Прекращение вегетации определяется сроками наступления осеннего похолодания – и амплитуда этой фенофазы снова увеличивается до месяца. Таким образом, в условиях интродукционного пункта I. рseudacorus проходит цикл сезонного развития в полном объеме.
Реальная семенная продуктивность (РСП) зрелых генеративных особей I. рseudacorus (табл. 2) позволяет размножать этот вид семенным путем и получать сеянцы практически в неограниченных количествах: РСП одного побега составляет более 500 семян, общее же количество цветоносов определяется прежде всего возрастом особи и составляет от одного до 20.
Таблица 2. Реальная семенная продуктивность (РСП) Iris pseudacorus L. в условиях Донецкого ботанического сада.

Примечание: M±m – средняя величина ± ошибка среднего; CV % – коэффициент вариации.

Рис. 2. Полиморфизм семян Iris pseudacorus L.
Для семян I. рseudacorus характерен некоторый полиморфизм: в большинстве своем они полукруглые или округлые, сплюснутые, однако небольшое количество семян, расположенных в верхней или нижней части плода, имеют коническую, овально-удлиненную или угловатую форму (рис. 2).
Однако в целом размеры семян характеризуются низким уровнем варьирования: коэффициент вариации не превышает 11 % (табл. 3). Размерные характеристики плода также характеризуются низким и средним уровнями варьирования, свидетельствующими о стабильном состояниии генеративной сферы интродуцента, что также подтверждает высокий уровень адаптации вида к новым условиям произрастания.
Таблица 3. Размерные характеристики плодов и семян Iris pseudacorus L. в условиях Донецкого ботанического сада.
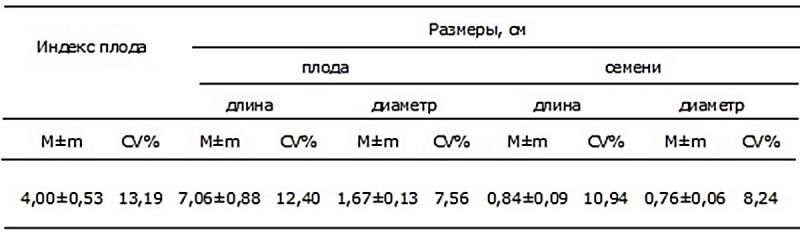
Примечание: M±m – средняя величина ± ошибка среднего; CV % – коэффициент вариации.
Варьирование РСП как особи, так и элементарной единицы, а также размеров коробочки I. pseudacorus определяется не только возрастным состоянием растения и погодными условиями в разные годы. При сравнении наших данных с аналогичными данными, полученными при интродукции I. pseudacorus в Уфимском ботаническом саду Республики Башкортостан (УБС) (Крюкова, Абрамова, 2014; Шайбаков, Миронова, 2008) очевидно определяющее значение и природно-климатических условий интродукционного пункта. Несмотря на то, что в обоих случаях интродуценты попадают в более засушливые условия степной зоны, при интродукции в ДБС отмечены значительно более высокие семенная продуктивность и морфометрические показатели плодов. Так, в нашем случае РСП плода почти в 2 раза превышает этот показатель в условиях УБС: 55,2 и 28,8 соответственно. Длина и ширина плода при интродукции составили 7,1 и 1,7 см против 4,1 и 1,3 см. Вероятно, не менее важную роль, чем режим увлажнения и температурный режим, для растений-интродуцентов играют эдафические условия интродукционного пункта, поскольку именно по этому признаку наиболее различаются условия ДБС и УБС (обыкновенные черноземы и серые лесные почвы).
Семена I. рseudacorus характеризуются пролонгированным периодом органического покоя: при весеннем посеве всходы появляются через 40–50 дней. Для изучения онтогенеза семена были высеяны осенью в неотапливаемой теплице, в конце мая следующего года сеянцы высажены в открытый грунт. На протяжении всего развития их измеряли, зарисовывали, фотографировали. В течение 14 лет растения последовательно прошли следующие возрастные состояния: проросток, ювенильное (первый год развития), имматурное (2-й год), виргинильное (3-й год), раннее генеративное (4–5-й годы), зрелое генеративное (5–6-й годы), позднее генеративное (10–11-й годы), субсенильное (12–13-й годы).
Некоторый полиморфизм семян I. рseudacorus (форма, размеры, толщина семенной кожуры и пр.), видимо, обусловливает растянутый период их прорастания (около 10 дней). Наличие этого периода предопределяет динамическую поливариантность всего онтогенеза, проявляющуюся в неодновременном достижении сеянцами очередного возрастного состояния. В результате 5–7 % особей отличаются замедлением смены возрастных состояний и около 2 % – ее ускорением.
Прегенеративный период
Проростки. При осеннем посеве всходы появляются во второй половине апреля следующего года. Это возрастное состояние наиболее динамично, структура проростков очень быстро усложняется, а потому можно выделить ранние и поздние проростки (рис. 3).

Рис. 3. Развитие проростка Iris pseudacorus L.
На начальной стадии развития проросток питается только за счет эндосперма семени и состоит из главного корня, бесцветного колеоптиля и первичного листа длиной в несколько мм. Тип прорастания гипогеальный: гипокотиль недоразвивается, семядоля остается под землей, а первичный лист высотой 3–5 см, приобретающий мечевидную форму и зеленую окраску, выносится на поверхность почвы. Затем появляются второй и третий листья, формируя характерный для ирисов вегетативный побег-веер высотой 9–10 см. Питание проростка становится смешанным, осуществляясь за счет эндосперма семени и ассимиляционных процессов, происходящих в листьях. Главный корень по мере роста ветвится до второго порядка, развиваются 2–4 придаточных корня, достигая 2,5–3,5 см длины. Это возрастное состояние длится 25–35 дней.
Г. И. Родионенко (1961) обращал внимание на свойственную проросткам I. pseudacorus ярко выраженную антигеофилию – выдвижение почечки в оптимальные для ее развития горизонты почвы, особенно ярко проявляющуюся при излишне глубокой заделке семян. Такую особенность прорастания семян он связывает с переувлажненными и слабо аэрируемыми грунтами в местах естественного произрастания вида. В нашем же случае, в условиях частого пересыхания верхних слоев почвы уже в мае и даже конце апреля, эту особенность I. pseudacorus можно считать важной адаптационной стратегией вида, дающей возможность проросткам благополучно развиваться в засушливых условиях степной зоны: более глубокая заделка семян при посеве создает проросткам оптимальные условия увлажнения, повышая их жизнестойкость.
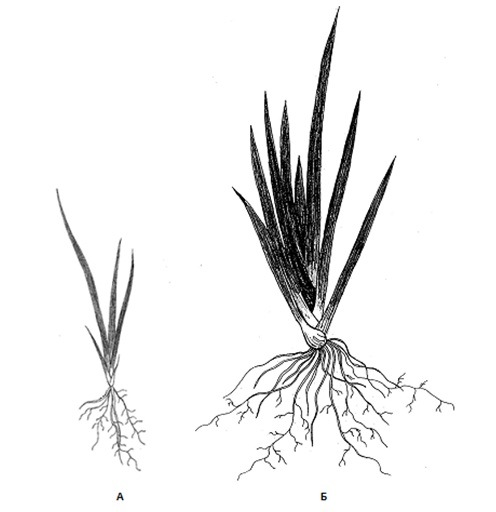
Рис. 4. Ювенильная (А) и имматурная (Б) особи Iris pseudacorus L.
Ювенильное возрастное состояние (рис. 4, А) характеризуется переходом к самостоятельному питанию и потерей связи с семенем. Листья увеличиваются в размерах до 12–16 см, их количество возрастает до 4, главный корень замедляет рост, а придаточные и боковые удлиняются. Количество придаточных корней возрастает до 5–7. У 10–15 % особей физическая связь с семенем продолжает сохраняться. Это возрастное состояние продолжается до конца вегетационного периода первого года развития. В конце лета базальная часть неветвящихся корней становится поперечно-морщинистой и они превращаются в контрактильные, втягивая формирующееся корневище глубже в почву. С наступлением холодов листья засыхают, их влагалища защищают почку возобновления в зимний период. В таком состоянии растение зимует.
В имматурное возрастное состояние сеянцы переходят на второй год развития. Сохраняется моноподиальное нарастание побега, в течение вегетационного периода развивается 7–8 мечевидных листьев длиной от 33 до 48 см, шириной 1,9–2,4 см. Подземная часть представлена коротким корневищем с множеством придаточных корней (рис. 3, Б). Половина из них разветвленные, более длинные (12–15 см длиной), остальные более толстые и короткие (до 10 см), шнуровидные. В конце лета шнуровидные корни превращаются в контрактильные, листья в конце октября сначала желтеют, затем засыхают.
Виргинильное возрастное состояние. Отличительная особенность этого возрастного состояния – становление жизненной формы в результате перехода от моноподиального к симподиальному нарастанию побегов с образованием первичного куста (рис. 5). На третьем году развития в результате ветвления корневища формируются особи с двумя или четырьмя вегетативными побегами. Каждый побег состоит из 8–12 срединных мечевидных листьев длиной 40–70 см, шириной 2,2–2,8 см. Подземная часть представлена коротким корневищем с множеством придаточных корней. Половина из них разветвленные, более длинные (до 15 см длиной), остальные более короткие и толстые (до 10 см длиной, 2–4 мм толщиной), шнуровидные, в конце лета превращающиеся в контрактильные. В этом году завершается прегенеративный период онтогенеза I. рseudacorus.

Рис. 5. Виргинильная особь Iris pseudacorus L.
Генеративный период
На четвертом году развития растения переходят в раннее генеративное возрастное состояние. Корневище продолжает ветвиться, в результате чего формируются особи с 6–8 вегетативными и одним генеративным побегом (рис. 6, А). Корневище хорошо выражено, 1,6–2,2 см толщиной, на 2,0–2,5 см от точки нарастания покрыто бурыми волокнами – остатками отмерших листьев генерации предыдущего вегетационного периода. Вегетативный побег состоит из 6–8 мечевидных листьев длиной 47–70 см, шириной 2,5–3,2 см. Генеративный побег высотой 50–58 см несет 4 ярко-желтых цветка 7 см диаметром, 4 см высотой. Из них только один цветок (в большинстве случаев верхний) развивается в коробочку с 16–25 полноценными семенами. В конце июля рост корней прекращается, значительная их часть становится контрактильными, втягивая корневище в почву. Вообще явление гетероризии (образование корней разных типов) у I. рseudacorus является сезонным и характерным для всех возрастных состояний, кроме проростков и ювенильных. Продолжительность этого возрастного состояния – один-два года.
Зрелое генеративное возрастное состояние. Особи характеризуются максимальным развитием вегетативной и генеративной сфер (рис. 6, Б). Растения мощные, побегообразование интенсивное, цветение обильное, плодоношение максимальное и регулярное. В этом возрастном состоянии растения пребывают 4–5 лет.

Рис. 6. Ранняя (А) и зрелая (Б) генеративные особи Iris pseudacorus L.
Позднее генеративное возрастное состояние (рис. 7, А). У отдельных особей с 9-го, а у большинства с 10-го года жизни начинаются процессы отмирания отдельных звеньев корневищ прошлых лет, сопровождающиеся снижением генеративной функции: количество генеративных побегов на особь снижается с 10–20 до 1–3. Продолжительность этого возрастного состояния составляет от 2-х до 4-х лет.
Постгенеративный период
Субсенильное возрастное состояние отмечено на 11–13-й годы развития и характеризуется в первую очередь отсутствием генеративных побегов, листья, сохраняя прежнюю длину, становятся значительно уже и поникают. В центральной части дерновины в результате отмирания и выпадения звеньев корневища отчетливо различим отмерший участок неправильной формы (рис. 7, Б).
В таком состоянии растения продолжают оставаться не менее 4–5 лет. Если корневище субсенильной особи разделить на небольшие фрагменты с 2–3 почками, нарастание вегетативной части интенсифицируется, происходит так называемое омоложение, в результате чего через виргинильное возрастное состояние растения возвращаются к раннему, а затем зрелому генеративному.

Рис. 7. Поздняя генеративная (А) и субсенильная (Б) особи Iris pseudacorus L.
Сенильный период
Сенильное возрастное состояние, характеризующееся крайней дряхлостью и резкой дегенерацией растений, по прошествии 15 лет нами пока не выявлено.
Выводы и заключение
Таким образом, нами установлены продолжительность прегенеративного периода онтогенеза I. рseudacorus в условиях культуры степной зоны (3 года) и периода декоративности особей (5–7 лет). Эти данные, совместно с данными о семенной продуктивности, позволят организовать процесс семенного размножения данного вида и прогнозировать долговременность существования экспозиций с его участием.
Для получения сравнительно небольшого количества посадочного материала I. рseudacorus целесообразно размножать вегетативно – делением корневищ виргинильных или генеративных особей по числу вееров ранней весной (апрель - начало мая) или более крупными фрагментами в период летне-осеннего покоя (август - сентябрь).
Оценка успешности интродукции в ДБС данного вида, гигромезофита и палюданта, доказывает обоснованность индивидуального подхода к проблемам введения дикорастущих видов в культуру: I. pseudacorus получил максимальную оценку (7 баллов): при регулярном поливе особи этого вида отличаются высокой устойчивостью к местным климатическим условиям, ежегодно массово цветут и плодоносят, активно разрастаются и дают самосев (рис. 8).
Полное отсутствие полива вызывает некоторое снижение морфометрических показателей и почти исключает самосев, однако цветение и созревание семян и в этом случае происходит ежегодно.
На основании проведенных исследований, I. рseudacorus нами рекомендуется как устойчивое и неприхотливое растение для широкого использования в зеленом строительстве Донбасса – для водоемов, миксбордеров, рокариев и гравийных садов, а также для срезки. В экспозициях ландшафтного типа он хорошо сочетается с нивяником, дербенником иволистным, ирисом сибирским, мелколепестником крупноцветковым, колокольчиком средним, лихнисом халцедонским, маком восточным и другими высокими многолетниками.

Рис. 8. Развитие самосева Iris pseudacorus L. в Донецком ботаническом саду.
Литература
Баканова В. В. Цветочно-декоративные многолетники открытого грунта. Киев: Наукова думка, 1984. 155 с.
Безделев A. Б., Безделева Т. А. Жизненные формы семенных растений российского Дальнего Востока. Владивосток: Дальнаука, 2006. 296 с.
Борисова И. В. Сезонная динамика растительного сообщества // Полевая геоботаника. Т. 4. Л.: Наука, 1972. С. 5—136.
Вайнагий И. В. О методике изучения семенной продуктивности растений // Ботан. журнал. 1974. Т. 59. № 6. С. 826—831.
Игнатьева И. П. Онтогенетический морфогенез вегетативних органов травянистых растений. М., 1983. 55 с.
Крюкова А. В., Абрамова Л. М. К биологии редкого вида Республики Башкортостан Iris pseudacorus L. в природе и интродукции // Научные ведомости Белгородского государственного ун-та. Сер. Естественные науки. 2014. № 17 (188). Вып. 28. С. 32—34.
Методика фенологических наблюдений в ботанических садах СССР. М., 1975. 42 с.
Работнов Т. А. Определение возрастного состава популяций видов в сообществе // Полевая геоботаника. Т. 3. М. - Л.: Наука, 1964. С. 132—208.
Работнов Т. А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Тр. БИН АН СССР. 1965. Сер. 3. № 6. С. 7—204.
Родионенко Г. И. Род Ирис – Iris L. (вопросы морфологии, биологии, эволюции и систематики). М. – Л.: Издательство Академии наук СССР, 1961. 216 с.
Тарасов В. В. Флора Днiпропетровської та Запорiзької областей. Донецьк: Лiра, 2012. 296 с.
Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени и энергетических процессов // Биол. науки. 1975. № 2. С. 7—34.
Федченко Б. А. Род 307. Касатик – Iris // Флора СССР. В 30-ти томах. М. – Л.: Издательство Академии наук СССР, 1935. Т. IV. С. 518.
Шайбаков А. Ф., Миронова Л. Н. Опыт интродукции редких видов ириса флоры Башкортостана // Бюл. бот. сада Саратовского государственного ун-та. Биология. 2008. С. 175—182.
Энциклопедия декоративных садовых растений; http://flower.onego.ru/voda/iris_pse.html (дата обращения 02.03.2017).
